Ualmindelig lav termisk modenhed af lagene
Thermisk modenhed er den vigtigste enkeltfaktor, der påvirker bevarelsen af gamle sedimentære biomarkører, da organiske molekyler er modtagelige for strukturelle og stereokemiske ændringer under gradvis nedgravning og med stigende termisk stress29. Prækambriske bjergarter med passende termisk modenhed (olievinduets modenhed eller lavere) er en forudsætning for at bevare robuste og primære biomarkørlipidsamlinger. Flere stereoisomerforhold mellem hopan- og steranbiomarkører samt uafhængige beviser fra lave Tmax-værdier (for det meste inden for et interval på 417-433 °C, med et gennemsnit på 426 °C) fra Rock-Eval-pyrolyse viser, at klipperne i denne undersøgelse repræsenterer langt de mest termisk umodne Ediacaran-sten, der er analyseret til dato ved hjælp af de mest avancerede organisk-geokemiske metoder (Tabel 1). Selv om vores bjergarter har gennemgået en sedimentær diagenese, har de ikke passeret betydeligt ind i olievinduet bortset fra de lidt mere modne 16PL-prøver fra Podillya-bækkenet i Moldova, som stadig er velegnede til analyse (tidlig til middelmådig olievinduets modenhed og ingen tydelige tegn på organiske forureninger). Størstedelen af sættet havde da sandsynligvis ikke været udsat for temperaturer i graven på over 50 °C, og vores prøver er derfor meget umodne. Andre linjer af molekylære beviser, som understøtter lav termisk modenhed og syngenicitet, omfatter (i) en dominans af polycykliske biomarkøralkaner frem for n-alkaner i bjergartsekstrakter (Fig. 2), (ii) overlevelse af påviselige mængder af 17β,21β(H)-hopaner, der kan opløses fra de mere rigelige hopaner, der besidder stabile 17β,21α(H)- og 17α,21β(H)-stereokemiske konfigurationer (Tabel 1, Supplerende figurer. 1 og 2), (iii) en tydelig odd-over-even præference blandt n-alkanerne i C22 til C27-området (Fig. 2), da n-alkaner kun viser en kulstofnummerpræference før katagenese38 , og (iv) generering af termisk umodne hopan- og steranbiomarkører fra den (uopløselige) kerogenfase ved hjælp af katalytisk hydropyrolyse (Supplerende Fig. 2). Den lave termiske modenhed af sedimentært organisk materiale i vores prøver er i overensstemmelse med tidligere offentliggjorte indikatorer for sedimentær alteration, herunder conodont- og acritarch-alterationsindeks, og tidligere Rock-Eval pyrolyse- og biomarkørundersøgelser af Ediacaran og Paleozoiske sedimentære bjergarter i Baltica36,39,40, samt lermineralogi41. Det er kritisk, at i omgivelser, hvor den termiske modenhed er så lav, er mekanismerne for betydelig ændring af primære δ15N- og δ13C-signaler stort set fraværende42.

Fordeling af ekstraherbare alifatiske kulbrinter for en repræsentativ prøve. a Totalionkromatogram (TIC) for ekstraherbare alifatiske kulbrinter for Lugovoe #13-73 m fra Redkino Horizon. N-alkan-serien, pristan (Pr), phytan (Ph) og C27-C34-hopaner (angivet ved deres samlede kulstofantal og stereokemi ved C-17, C-21 og C-22, f.eks. C31αβR) er mærket; ** angiver C29- og C30-hopener. Bemærk hopanernes dominans i forhold til andre klasser af alkanforbindelser. b Delvis 85-Da ionkromatogram viser dominans af n-alkan-serien, der udviser en tydelig præference for ulige over lige kulstofnumre i C22-C27-området, over methylalkaner
Lipid biomarker assemblage mønstre
Et abundansforhold mellem de vigtigste (C27-C35) hopaner og de vigtigste (C27-C29) steraner bruges ofte til at vurdere balancen mellem bakterielle versus eukaryote kildeorganismers input til det gamle akvatiske økosystem. Hopaner er molekylære fossiler, der stammer fra hopanoider, som er cellemembranlipider, der syntetiseres af en lang række forskellige bakteriegrupper. På samme måde er steraner afledt af sterolprækursorer, som næsten udelukkende produceres af eukaryoter43. Hopan/steran (H/St)-forholdet for vores prøver dækker et påfaldende stort interval af værdier fra 1,6 til 119,2 (tabel 1). Til sammenligning falder H/St-forholdet fra organisk rige neoproterozoiske bjergarter og olier typisk i et snævert interval fra 0,5 til 2,030,31,32,33. Selv om der er en klar forskel på værdierne fra den yngre Kotlin Horizon (gennemsnitlig H/St på 8,9) og den ældre Redkino Horizon (gennemsnitlig H/St på 42,9), er værdierne for de fleste prøver usædvanligt høje og tyder på unormalt høje bidrag fra bakterier.
I slutningen af Neoproterozoikum var eukaryotiske alger en økologisk betydningsfuld komponent og store producenter i mange marine økosystemer15,34,44. Diskrepansen mellem det globalt vigtige bidrag fra eukaryotiske alger i det sene neoproterozoikum og alligevel de lave niveauer af steranbiomarkører i disse prøver tyder på, at der må være en eller anden lokal determinant for eukaryotiske forekomster. Den mest nærliggende forklaring på de ekstremt forhøjede hopan/steran-forhold sammen med et lavt indhold af organisk kulstof (TOC) og et lavt hydrogenindeks (HI) i vores prøver (tabel 1) er, at disse lag blev aflejret i oligotrofe (dvs. stærkt næringsstofbegrænsede) omgivelser, hvor bakterier udkonkurrerede alger. Mens moderne analoger for gamle epikratoniske have udviklet under høj havniveaustand er svære at finde, er der tidligere fundet parallelle observationer af højere hopan/steran-forhold (med op til en størrelsesorden) for organisk magre kontra organisk rige sedimentære bjergarter aflejret i Ordovicium-Siluriske epikontinentale havveje45.
Typisk kan et lavt TOC-indhold skyldes begrænset aflejring af organisk stof i lavproduktive omgivelser, lavt bevaringspotentiale af organisk stof i vandsøjlen eller sedimenterne eller fortynding med en høj siliklastisk flux. I moderne oceaner er produktiviteten oftest begrænset af lave niveauer af de essentielle næringsstoffer: kvælstof, fosfor, silicium og jern46 . Kvælstof, fosfor og jern er begrænset i områder i det åbne hav, hvor opstrømning, støvtilførsel og afstrømning fra kysterne ikke tilfører tilstrækkelige mængder næringsstoffer. I moderne oligotrofe miljøer er forholdet mellem bakteriel og eukaryotisk biomasse højere end i eutrofe eller mesotrofe miljøer. Det er interessant, at Podillya 16PL-udskæringsprøverne fra Redkino-horisonten i Moldova er fosforitholdige muddersten og giver betydeligt lavere hopan/steran-forhold (8,1:11,5) end de andre Redkino-prøver i tabel 1 (dog stadig betydeligt højere end dem, der er fundet i South Oman Salt Basin eller i organisk-rige Phanerozoiske sedimenter, som typisk falder inden for et snævert, 0.5-2,0 interval), hvilket muligvis tyder på, at øget fosfattilgængelighed kan have haft en gunstig indflydelse på det eukaryote-til-bakterie-forhold, der er fundet lokalt (i betragtning af at kvælstofisotopsignaturerne stort set er invariante, se næste afsnit). Fosforindholdet (P) samt P/Fetotal og P/Al-forholdet er ellers generelt lavt for de sene ediakariske sedimenter fra Podillya, Ukraine og Estland47; med undtagelse af dette stratigrafiske niveau, der er markeret med fosforitknuder. Tilsvarende blev der rapporteret om lave (~0,01 til ~0,1 vægtprocent) niveauer af P i Kotlin og Redkino silikoklastiske bjergarter fra en borekerne fra den nordøstlige margin af den østeuropæiske platform11.
Den brede og lavvandede topografi i de epikontinentale have på tværs af Baltica kunne have opretholdt fosfor- eller andre næringsstofbegrænsninger i marginale omgivelser, som følge af authigen udfældning af fosfat med jernmineraler i det oxiske overfladevand og fastholdelse af en række grundstoffer i shelfsedimenter48. Hvis fosfat var et begrænsende næringsstof, ville øget biotilgængeligt fosfor have øget den lokale primærproduktion og skabt mere gunstige vækstbetingelser for større encellede fytoplankton49,50,51. Med hensyn til det moderne havsystem er det kendt, at de marine picocyanobakterier, Prochlorococcus og Synechococcus, dominerer antallet af fytoplanktonceller og biomasse i oligotrofe tropiske og subtropiske havmiljøer, herunder fosfatbegrænsede oligotrofe områder i havets overfladevand52. Prochlorococcus og den heterotrofe SAR11 (Pelagibacter) trives på grund af en række tilpasninger, herunder lave energiomkostninger i kraft af små genomer og lave replikationshastigheder, et højere overfladeareal/volumen-forhold i kraft af mindre cellestørrelser og yderligere celleoptagelsesfunktioner for at maksimere udnyttelsen af næringsstoffer53. Mange bakterier er også i stand til at erstatte næringsstoffer med lavt indhold, f.eks. ved at udnytte sulfolipider i stedet for fosforholdige lipider i P-manglende omgivelser54 , eller ved at anvende alternative substrater, f.eks. ved at hente kvælstof fra atmosfærisk N2 via nitrogenfiksering55 , for at afhjælpe næringsstofstress. En nylig undersøgelse af marine bakterioplanktons overlevelse i oligotrofe miljøer56 , hvor der kun er begrænset fosfat til rådighed, tyder på, at polyfosfatmetabolismen spiller en vigtig rolle for marine oligotrofe organismer. De udvalgte eukaryoter, der konkurrerer i oligotrofe miljøer, er typisk små picoeukaryoter, som kan supplere deres ernæringsbehov gennem mixotrofi57. Forholdet mellem picoeukaryoter og cyanobakteriernes biomasse har en tendens til at stige under øget næringsstoftilførsel49. Den mulige indflydelse af fosfor og andre biolimiterende næringsstoffer på modereringen af primærproduktiviteten og den marine samfundsstruktur i epeiriske havområder fra Ediacaran kræver yderligere undersøgelser.
Op alle vores lokaliteter (tabel 1) er forekomsten af C29-steraner uden undtagelse større end de tilsvarende C27- eller C28-steraner. En overvægt af C29-steraner i forhold til C27- og C28-steraner indikerer sandsynligvis en grønalgedominans inden for det eukaryote fytoplanktonsamfund44,58. Dette træk er blevet observeret i de fleste tidligere Ediacaran biomarkørundersøgelser15,30,31,32,32,33,34,35,36,44. Det er bemærkelsesværdigt, at C30-steranfordelingen i flere prøver fra hver borekerne i Kotlin Horizon indeholder lave, men påviselige mængder af demosponge-steranbiomarkøren kendt som 24-isopropylcholestan (24-ipc)59,60. I alt er der rapporteret om 24-ipc steraner i sten- og olieprøver, der er dateret så langt tilbage som Cryogenian (>635 Ma siden) i South Oman Salt Basin og repræsenterer det ældste lipidbiomarkørbevis for metazoer59,60. 24-ipc-biomarkørerne i vores prøver var enten omkring en størrelsesorden lavere i overflod (i forhold til de samlede C27 til C30 steranforhold var disse kun 0,06-0,61 %; gennemsnit = 0,22 %) sammenlignet med Ediacaran klipper og olier fra Sydomans (1,7 % i gennemsnit59) eller var under detektionsgrænserne på grund af ubetydelig overflod for størstedelen af prøverne.
Stikstof- og organisk kulstofisotopforhold
Stikstofisotoper kan hjælpe med at skelne den relative balance i kvælstofkredsløbet, og i hvilken grad enten kvælstoffiksering eller ufuldstændig denitrifikation var væsentlige veje til at påvirke den næringsstofbalance, der var tilgængelig for marine samfund. Når diazotrofe bakterier fikserer molekylært kvælstof på grund af mangel på fikseret kvælstof i vandsøjlen, kan dette give sedimentære bulk-kvælstofisotopværdier nær 0‰61. Dette står i kontrast til de positive kvælstofisotopsignaturer (i intervallet +2 til +10‰) med en modus på +4 til +6‰, der er fundet for de neoproterozoiske marine sedimenter, der er aflejret under, hvad der menes at være nitratrige forhold, hvor nitrat kun har undergået delvis denitrifikation62. Kvælstofisotopværdierne for alle vores prøver bortset fra en enkelt udbryder (Tabel 1) dækker et begrænset positivt område fra +3,5 til +6,5‰, som overlapper med modus for sene neoproterozoiske sedimentære bjergarter, der er rige på organisk materiale62. Begrænset af redoxproxies, der tyder på, at der herskede oxiske vandsøjleforhold, antyder vores data, at nitrat dominerede den opløste uorganiske kvælstofpulje. Spektret af δ15N-værdier tyder også på, at N2-fiksering ikke var den primære måde for primærproducenterne at erhverve kvælstof på. I stedet var kvælstofkredsløbet sandsynligvis domineret af genanvendelse i vandsøjlen, og organisk N var genstand for kvantitativ oxidation til nitrat, som det er tilfældet i moderne, proximale marine miljøer. Benthisk denitrifikation og nedgravning af organisk N ville have været de primære dræn for opløst uorganisk kvælstof, og intervallet af δ15N-værdier tyder kun på en begrænset rolle for ufuldstændig denitrifikation i vandsøjlen, som typisk resulterer i en betydelig 15N-berigelse63. Den begrænsede variabilitet i δ15N-værdierne afspejler derfor sandsynligvis δ15N af nitrat, der er advekteret til platformen, med mindre indflydelse fra begrænset nitratreduktion og N2-fiksering i vandsøjlen. Uden stærke δ15N-beviser for kvælstoffiksering antager vi, at kvælstof ikke var det primære biolimiterende næringsstof i de epikontinentale bassiner i Baltica. Isolation fra flod- og eoliske fosforkilder og oxiske forhold i de brede, lavvandede, marine epikontinentale bassiner kan have forbedret fjernelsen af authigenisk fosforit og spormetalholdige faser, hvilket resulterede i en næringsstofbegrænsning, der begrænsede eukaryotisk cellevækst og produktion.
Total organisk kulstof (CTOC) isotopværdier varierer fra -23,0 til -33,9‰, med de største forskelle observeret mellem de forskellige borekernelokaliteter. Den relative 13C-berigelse til isotopforhold højere end ca. -28‰ står i kontrast til data fra samtidige lag aflejret i eutrofe, åben-marine miljøer i Oman30,31. Det interval, vi rapporterer, er dog generelt i overensstemmelse med δ13CTOC-området for andre lokaliteter fra Baltica, der er aflejret i samme periode11. Forskellen mellem Baltica og Oman kan understrege det faktum, at δ13CTOC-værdierne ikke udelukkende afspejler en ensartet sekulær ændring i kulstofkredsløbet i dette tidsinterval31,64. Snarere kan δ13CTOC-området fra Baltica til dels afspejle den bakterielt dominerede mikrobielle økologi, som de unikke lipidbiomarkørforhold tyder på. Små cellestørrelser, høje overfladeareal-til-volumen-forhold og langsomme vækstrater under oligotrofe forhold kan øge størrelsen af fraktioneringen under autotrofi (εp)65,66, hvilket resulterer i lave δ13C-værdier.
Mekanismen for de mere 13C-berigede isotopiske signaturer inden for det område, der er rapporteret11 for den sene Ediacaran Baltica-succession, er mindre klar. Det potentielle bidrag fra detritalt, metamorfisk ændret organisk materiale til prækambriske sedimentære successioner med lavt TOC-indhold er en mulig mekanisme for forskelle mellem lokaliteterne i δ13CTOC64. Imidlertid er TOC-indholdet i vores prøver, selv om det er lavt, generelt større end 0,10 vægtprocent, hvilket ikke afslører nogen sammenhæng mellem TOC-indholdet og δ13C-værdierne. Desuden indeholder vores prøver termisk umodent organisk materiale og biomarkørlipidmønstre, som ikke stemmer overens med en hovedsageligt allochton kulstofkilde. Mere 13C-berigede δ13CTOC-signaturer kan være resultatet af alternative mekanismer for kulstofassimilation. Kulstofkoncentrationsmekanismer eller prokaryoternes aktive bicarbonatoptagelse kan resultere i mindre værdier for εp og højere δ13CTOC-værdier67,68. Fremkomsten af den ediacariske biota kan have udvidet de marine fødekæder betydeligt og stimuleret nye muligheder for mikrobiel heterotrofi, herunder mulige bidrag fra komplekse kulstofcyklusser i bentiske mikrobielle måtter. Med yderligere hensyntagen til den potentielt vigtige rolle, som opløst organisk stof69 spiller som kulstofkilde, afspejler bredden af δ13CTOC-værdierne sandsynligvis en række biogeokemiske kulstofcyklusprocesser, der kan være unikke for det udviklende ediacariske havmiljø og kan være relateret til et fænomen, som vi ikke har nogen repræsentative moderne analoger for.
Man har fundet gamle samlinger af lipidbiomarkører og stabile isotopindikatorer for bentisk, mikrobiel matteproduktion i forbindelse med fast kvælstofbegrænsning i de tidlige triasiske bjergarter fra Sydkina70. I kølvandet på masseudryddelsen i slutningen af Perm ved Meishan var der en stor stigning i hopan/steran-forholdet (op til ca. 60) ledsaget af et stærkt skift i Norg-isotop-signaturen til værdier på 0 til -2‰, hvilket stemmer overens med bakteriel diazotrofi. Dette er forbundet med et højt signal af 2-methylhopaner (2-methylhopanindeks på op til 33 %) og karakteristiske methylalkaner, som peger på spredning af bentiske mikrobielle måtter. Vi observerer ikke en lignende tendens i kvælstofisotopsignatur eller biomarkørmønstre, som tydeligvis ville pege på et betydeligt mikrobielt matteinput i vores Ediacaran-datasæt, selv om der blev fundet en enkeltstående 15N-forarmet værdi i vores prøver (Tabel 1). Derfor er det stærke bakterielle signal, der er observeret i vores Baltica-prøver, sandsynligvis ikke overvejende en signatur af bentiske mikrobielle måtter.
Palæomiljømæssig næring af Ediacara-biota vs. demosponges
De tilsyneladende oligotrofe forhold på tværs af de epikratoniske og kontinentale randbassiner i Baltica, da det drev fra høje til lave breddegrader fra den sene Ediacaran til tidlig Kambrium, kan være forbundet med enten begrænset advektion af relativt næringsrige dybe vande eller udtømning af næringsstoffer, som følge af assimilation og scavenging under transport og aflejring på tværs af disse brede, lavvandede epikontinentale bassiner (Fig. 3). Disse bassiner blev episodisk isoleret fra havene og udviklede hypersalinitet (f.eks. i Redkino-tiden) og brakvandsforhold (f.eks. i Kotlin-tiden71). Langvarig tektonisk stabilitet resulterede i en topografi med lavt relief i det sene Ediacaran Baltica, som var meget modtagelig for oversvømmelser og ineffektiv tilførsel af fosfor fra forvitring. I modsætning til Baltica afslørede biomarkørundersøgelser af lagene fra Huqf Supergroup i Oman et eutrofisk økosystem, der var rigt på mikroalger30,31,59, men som manglede Ediacara-biota, selv i omfattende udgravninger i Omanbjergene og Huqf-regionen, i de indre til ydre skelfylder3. Palæogeografien af det sydlige Oman-saltbækken er blevet rekonstrueret for det sene neoproterozoikum på ~13° fra ækvator på den sydlige halvkugle, hvilket stort set svarer til Baltica72-palæolatiteten.
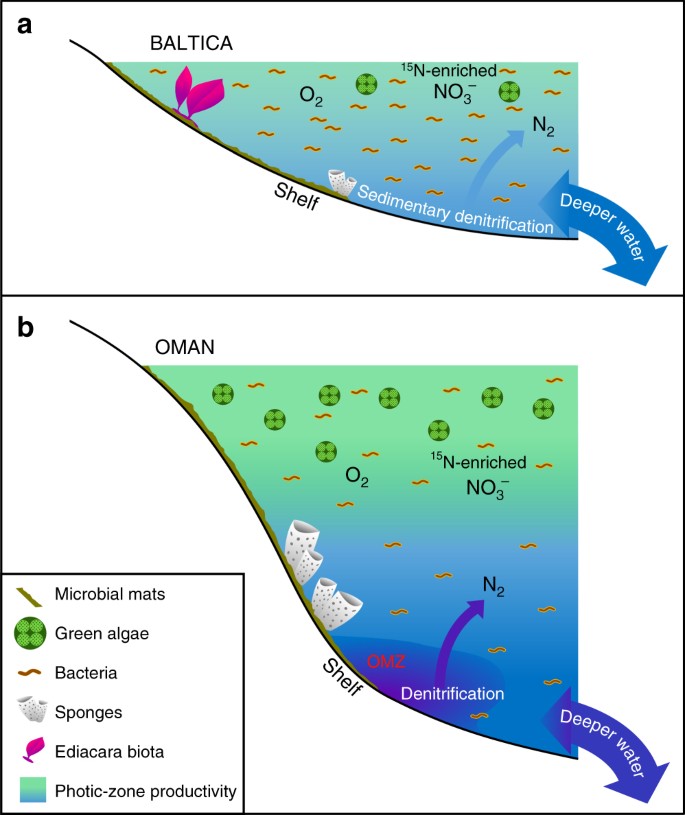
Større forskelle i lavproduktive vs. produktive Ediacaran havmiljøer. Der vises skematiske diagrammer for et omfattende oligotrofisk og lavvandet marint epikontinentalt bassin i Baltica, der ofte var domineret af bakteriel produktivitet, hvor Ediacara bløddyrsfaunaen blomstrede, og hvor denitrifikation og anammox sandsynligvis var begrænset til sedimenterne; og b eutrofe og dybere marine skelfiljøer i det sydlige Oman-saltbækken, hvor grønalger trivedes som primærproducent og demosponger var talrige, men hvor Ediacara-bløddyrsfaunaen ikke var fremtrædende (med Ediacara-biota-fossiler også fraværende i korrelative Ediacaran-fremspring i det nordlige Oman). På produktive kontinentalrande fandt denitrifikation og anammox sandsynligvis sted i både vandsøjlen og sedimenterne
Der er ingen beviser for vedvarende og omfattende anoxi på og omkring de lavvandede kontinentalrande i Baltica i løbet af det sene Ediacaran11, og vi foreslår, at de oligotrofe forhold, der er beskrevet her, blev forårsaget af ineffektive terrestriske og dybvands næringsstofstrømme til disse omgivelser, der i store træk svarer til dem i oligotrofe miljøer i moderne havsystemer. Faktisk tyder en geokemisk sporelementundersøgelse af Utkina Zavod og tilstødende borekerner fra St. Petersborg-området på, at vores prøver blev aflejret under oxiske forhold73. Denne fortolkning understøttes uafhængigt af vores data fra de udvidede hopan (C31-C35)-fordelinger, der aftager kraftigt i abundans over C31-forbindelser med stigende kulstofantal, hvilket er karakteristisk for sidekædenedbrydning af bakteriohopanepolyoler under diagenese under oxiske forhold29, og også ved lavt hydrogenindeks, som målt ved Rock-Eval pyrolyse (alle lavere end 230 mg/g TOC, og mange er under 100 mg/g TOC) for disse umodne prøver som følge af dannelsen af recalcitrant kerogen via oxidativ nedbrydning og rekondensering af lipidfattig primær biomasse i lokalt oxiske miljøer (Tabel 1). De Ediacariske flercellede organismer, der eksisterede i disse lavvandede havmiljøer, må have haft tilstrækkelige organiske substrater til heterotrofi til at opfylde deres fødebehov sammen med tilstrækkelig ilt og andre næringsstoffer til at opretholde deres stofskifte. De epikontinentale bassiner i Baltica var sandsynligvis mere vedvarende oxiske end de meget produktive miljøer, der omkransede iltminimumszoner, som f.eks. på den midterste til yderste hylde i South Oman Salt Basin, hvor respiration af rigelig planktonbiomasse ville have opretholdt lavere opløst ilt under den photiske zone.
Ediacara biota og andre flercellede organismer, der lever ud for Baltica, ville have været nødt til at klare skiftende fødekilder (bakterielle vs. eukaryote), da næringsstofstrømme varierede gennem tiden, herunder små celler og organisk detritus i epikontinentale bassiner, hvor bakterier var de dominerende primærproducenter (Fig. 3). Moderne koralrevssamfund overlever i tropiske oligotrofe miljøer på grund af effektiv genanvendelse af næringsstoffer, herunder generering af opløst organisk stof (DOM), som er med til at opretholde heterotrofi af faunaen i revets økosystem74. Etableringen af en marine trofisk struktur med eukaryote flercellede organismer, der ernærer sig af organisk detritus, må have fundet sted efter den globale miljømæssige ekspansion af eukaryoter i forskellige havmiljøer, der fandt sted i Tonian-Cryogenian-intervallet (ca. 800-635 Ma), som det fremgår af biomarkørregistreringer15. Vi kan så forvente at finde beviser for betydelige forskelle i den marine samfunds- og trofiske struktur fra lokalitet til lokalitet i den sene Ediacaran-periode, hvor den lokale næringsstofbalance vælger eukaryote-rige eller eukaryote-arme mikrobielle samfund, og hvor den gradvise ekspansion af flercellede organismer tilføjer endnu en dimension af kompleksitet på organisme- og samfundsniveau. Betydelig regional kontrast i fosfat og anden næringsstoftilgængelighed i hyldemiljøer er også en forventet konsekvens af en heterogen global marin redoxstruktur i det sene Ediakaran, før iltningen af det dybe ocean38.
Dominansen af picoplankton bakteriel produktivitet og tilhørende nedbrydningsprodukter af opløst organisk stof (DOM) i Baltica epikontinentale bassiner kunne have opretholdt et mikrobielt loop-økosystem parallelt med den konventionelle trofiske struktur baseret på omkring større planktonproducenter50. Dette kan have begunstiget forskellige former for heterotrofi, herunder suspensionsfodring og muligvis osmotrofi, som en levedygtig fødestrategi for nogle rangeomorfer, svampe og andre flercellede organismer fra det sene Ediacaran18 sammen med den nye aktive (bevægelige) heterotrofi19,20. Næringsstofbegrænsede akvatiske systemer er som hovedregel ofte domineret af lille encelligt fytoplankton og heterotrofisk plankton, hvor bakterier udkonkurrerer eukaryoter og opretholder en lav nettobiomasse i oligotrofiske marine miljøer i det moderne hav49,51. DOM er en vigtig kilde til organiske næringsstoffer og styrer ofte produktiviteten og nettobiomassen i moderne oligotrofe tropiske have, men det samme gælder for kvælstof- og fosforkobling75. På samme måde var DOM sandsynligvis også et vigtigt substrat til opretholdelse af heterotrofe bakterier og et mikrobielt kredsløb i gamle oligotrofe miljøer. Bentiske mikrobielle måtter kan have været en del af dette bakteriedominerede fødekædenet, og måtter er blevet anset for at forbedre bevarelsen af Ediacara-biota i marine miljøer7. Mens der blev fundet moderate værdier af 2-methylhopanindekset for en delmængde af vores prøver (4-10 %, tabel 1), tyder meget lave forekomster af methylalkaner i forhold til n-alkaner (fig. 1) og kun sporadiske og sporadiske forekomster af carotenoider på, at mikrobielle måtter ikke dominerede den primære produktivitet76 , og at planktonbakterier og tilsvarende deres DOM-nedbrydningsprodukter blomstrede. De generelt lave hydrogenindeksværdier (Tabel 1; især for Redkino-prøverne), på trods af den lave termiske modenhed af lagene og biomarkørsamlinger, er også i overensstemmelse med lipidfattig organisk tilførsel i hovedsageligt oxiske, lavvandede marine marginale miljøer. Forekomsten af mikrobielle måtter, som kun vurderet ud fra sedimentologiske teksturer, fortæller os under ingen omstændigheder noget om balancen mellem eukaryoter (mikroalger) og bakterier, og de kan heller ikke begrænse det relative bidrag fra mikrobielt plankton til den samlede primære produktivitet og opretholdelsen af fødekæder. Mikrobielle mattsamfund, der ofte indeholder rigelige eukaryoter, og Ara-gruppens karbonater fra det sydlige Oman med fremtrædende thrombolitiske og krusede laminitfacies er et godt eksempel på sene neoproterozoiske miljøer med et betydeligt mikrobielt mattebidrag, der giver rigelige steransignaler på grund af et stort bidrag af mikroalger30,59.
Det ekstremt lave niveau af 24-ipc-biomarkører tyder på, at selv om demosponges undertiden var til stede, var de sparsomme i disse miljøer. Dette kunne indikere færre muligheder for mindre filterfødende dyr i konkurrence med Ediacara-biotaen i disse miljøer med lav produktivitet, utilstrækkelig modstandsdygtighed hos demosponges mod mere energirige lavvandede marine forhold over fair-weather bølgebasis eller bedre tilpasningsevne hos svampe til iltfattige forhold, der dynamisk opretholdes under den photiske zone i eutrofe miljøer (fig. 3). I alt blev 24-ipc ikke påvist i de fleste af prøverne fra Redkino Horizon, som også generelt har de højeste H/St-forhold blandt vores prøvesæt, hvilket måske indikerer en økologisk ændring ved Kotlin Horizon-aflejringstidspunktet. Alternativt er det også plausibelt, at svampe beboede disse miljøer, men ikke producerede disse diagnostiske steroidbiomarkører i rigelige mængder, selv om dette synes mindre sandsynligt, da 24-ipc steran er blandt de mest almindeligt påviste C30 steranforbindelser i Ediacaran-lag og -olier30,3133,59,60 og også kan påvises i en delmængde af vores prøver (Tabel 1).
Mens de vedvarende oligotrofe havmiljøer, der antydes af vores data, repræsenterer lokale forhold i de ediakariske oceaner, var de sandsynligvis ikke ualmindelige for prækambriske lavvandede havområder (Fig. 3). Langt størstedelen af tidligere Ediacaran biomarkørstudier er blevet udført på organisk-rige sedimentære bjergarter, der er aflejret i eutrofe miljøer og deres olieprodukter, som generelt giver biomarkørsamlinger, der er i overensstemmelse med et betydeligt bidrag fra mikroalger som kilde44. Vores resultater understreger vigtigheden af at studere en bredere vifte af aflejringsmiljøer, herunder lag af organisk-materielfattige lag af passende termisk modenhed og forskellige lithologier15,3034,59, for at få et mere præcist billede af omfanget af heterogenitet i marin kemi og økologi fra sted til sted. På trods af progressiv havventilation og øget kemisk forvitring og næringsstoftilførsel under Rodinias opløsning og i hele Ediacaran-perioden77,78 opretholdt havets heterogenitet en række forskellige marine kemiske forhold, herunder næringsfattige, men beboelige miljøer, der fremmede metazoernes tilpasning, konkurrence og evolution i det globale havsystem (Fig. 3). Mens eutrofe marine hylder var vært for demosponges, men ofte manglede Ediacara biota, med South Oman Salt Basin (og de korrelative tykke Ediacaran udgravninger længere nordpå i Oman) som et fremtrædende eksempel; kontraintuitivt blev lavvandede, oxiske og mindre produktive epikontinentale havveje koloniseret af Ediacara biota i stedet for demosponges på trods af DOM og andet organisk detritus, der var tilgængeligt lokalt til fodring. Det er fortsat uvist, om metaboliske krav eller miljømæssigt selektivt pres begrænsede Ediacaran blødkødede flercellede biota til disse omgivelser; vores undersøgelse fremhæver dog, at Ediacaran oligotrofe omgivelser spillede en potentielt afgørende rolle i udviklingen af makroskopiske flercellede organismer og marine samfundsøkologi.