Außergewöhnlich geringe thermische Reife der Schichten
Die thermische Reife ist der wichtigste Faktor, der die Erhaltung alter sedimentärer Biomarker beeinflusst, da organische Moleküle während des fortschreitenden Vergrabens und mit zunehmender thermischer Belastung strukturellen und stereochemischen Veränderungen unterliegen29. Präkambrische Gesteine mit angemessener thermischer Reife (Ölfensterreife oder niedriger) sind Voraussetzung für die Erhaltung robuster und primärer Biomarker-Lipidansammlungen. Mehrere Hopan- und Steran-Biomarker-Stereoisomer-Verhältnisse sowie unabhängige Hinweise auf niedrige Tmax-Werte (meist innerhalb eines Bereichs von 417-433 °C, mit einem Mittelwert von 426 °C) aus der Rock-Eval-Pyrolyse deuten darauf hin, dass die Gesteine in dieser Studie die bei weitem thermisch unreifsten Ediacaran-Gesteine darstellen, die bisher mit den modernsten organisch-geochemischen Methoden analysiert wurden (Tabelle 1). Obwohl unsere Gesteine eine sedimentäre Diagenese durchlaufen haben, sind sie nicht wesentlich in das Ölfenster eingetreten, abgesehen von den etwas reiferen 16PL-Aufschlussproben aus dem Podillya-Becken in Moldawien, die sich immer noch für die Analyse eignen (frühe bis mittlere Ölfenster-Reife und keine offensichtlichen Anzeichen für organische Verunreinigungen). Die Mehrzahl der Proben war wahrscheinlich noch nicht Temperaturen von über 50 °C ausgesetzt, so dass unsere Proben sehr unreif sind. Weitere molekulare Beweise, die für eine geringe thermische Reife und Syngenizität sprechen, sind (i) die Dominanz polyzyklischer Biomarker-Alkane gegenüber n-Alkanen in Gesteinsextrakten (Abb. 2), (ii) das Überleben nachweisbarer Mengen von 17β,21β(H)-Hopanen, die von den häufiger vorkommenden Hopanen mit stabilen 17β,21α(H)- und 17α,21β(H)-Stereochemie-Konfigurationen unterschieden werden können (Tabelle 1, ergänzende Abbildungen. 1 und 2), (iii) eine erkennbare ungerade-über-gerade-Präferenz unter den n-Alkanen im C22- bis C27-Bereich (Abb. 2), da n-Alkane nur vor der Katagenese eine Kohlenstoffzahl-Präferenz zeigen38, und (iv) die Erzeugung von thermisch unreifen Hopan- und Steran-Biomarkern aus der (unlöslichen) Kerogenphase durch katalytische Hydropyrolyse (ergänzende Abb. 2). Der geringe thermische Reifegrad der organischen Sedimentmaterie in unseren Proben steht im Einklang mit bereits veröffentlichten Indikatoren für die Alteration von Sedimenten, einschließlich Conodonten- und Akritarchen-Alterationsindizes, sowie mit früheren Rock-Eval-Pyrolyse- und Biomarker-Studien der Sedimentgesteine des Ediacaran und des Paläozoikums in Baltica36,39,40 und der Tonmineralogie41. Entscheidend ist, dass in Umgebungen, in denen die thermische Reife so gering ist, die Mechanismen für eine signifikante Veränderung der primären δ15N- und δ13C-Signale weitgehend fehlen42.

Verteilung der extrahierbaren aliphatischen Kohlenwasserstoffe für eine repräsentative Probe. a Gesamtionenchromatogramm (TIC) für extrahierbare aliphatische Kohlenwasserstoffe für Lugovoe #13-73 m aus dem Redkino-Horizont. Die n-Alkanreihe, Pristan (Pr), Phytan (Ph) und C27-C34-Hopane (bezeichnet durch ihre Gesamtkohlenstoffzahl und Stereochemie an C-17, C-21 und C-22, z. B. C31αβR) sind markiert; ** bezeichnet C29- und C30-Hopene. Man beachte die Dominanz der Hopfenhäufigkeit gegenüber anderen Alkanverbindungsklassen. b Das partielle 85-Da-Ionen-Chromatogramm zeigt die Dominanz der n-Alkanreihe, die eine erkennbare Bevorzugung ungerader gegenüber geraden Kohlenstoffzahlen im Bereich C22-C27 aufweist, gegenüber Methylalkanen
Lipid-Biomarker-Assemblage-Muster
Ein Häufigkeitsverhältnis zwischen den wichtigsten (C27-C35) Hopanen und den wichtigsten (C27-C29) Steranen wird häufig verwendet, um das Gleichgewicht zwischen bakteriellen und eukaryotischen Ursprungsorganismen in einem alten aquatischen Ökosystem zu bewerten. Hopane sind molekulare Fossilien, die sich von Hopanoiden ableiten, welche Zellmembranlipide sind, die von einer Vielzahl von Bakteriengruppen synthetisiert werden. In ähnlicher Weise leiten sich Sterane von Sterol-Vorläufern ab, die fast ausschließlich von Eukaryoten produziert werden43. Das Hopan/Steran-Verhältnis (H/St) unserer Proben umfasst eine auffallend große Bandbreite von Werten zwischen 1,6 und 119,2 (Tabelle 1). Zum Vergleich: Die H/St-Verhältnisse in organisch reichen Gesteinen und Ölen aus dem Neoproterozoikum liegen typischerweise in einem engen Bereich von 0,5 bis 2,030,31,32,33. Während es einen deutlichen Unterschied zwischen den Werten des jüngeren Kotlin-Horizonts (durchschnittliches H/St von 8,9) und des älteren Redkino-Horizonts (durchschnittliches H/St von 42,9) gibt, sind die Werte für die meisten Proben ungewöhnlich hoch und deuten auf einen anomal hohen Beitrag von Bakterien hin.
Im späten Neoproterozoikum waren eukaryotische Algen ein ökologisch bedeutender Bestandteil und Hauptproduzent in vielen marinen Ökosystemen15,34,44. Die Diskrepanz zwischen dem weltweit bedeutenden Beitrag eukaryontischer Algen im späten Neoproterozoikum und den geringen Mengen an Steran-Biomarkern in diesen Proben deutet darauf hin, dass es irgendeine lokale Determinante für die Häufigkeit von Eukaryonten geben muss. Die plausibelste Erklärung für die extrem hohen Hopan/Steran-Verhältnisse neben den niedrigen Gehalten an organischem Gesamtkohlenstoff (TOC) und den niedrigen Wasserstoffindizes (HI) in unseren Proben (Tabelle 1) ist, dass diese Schichten in einem oligotrophen (d. h. stark nährstoffbegrenzten) Umfeld abgelagert wurden, in dem Bakterien die Algen verdrängten. Während moderne Analoga für alte epikratonische Meere, die sich während eines hohen Meeresspiegelstandes entwickelten, schwer zu finden sind, wurden parallele Beobachtungen höherer Hopan/Steran-Verhältnisse (um bis zu einer Größenordnung) für organisch magere gegenüber organisch reichen Sedimentgesteinen gefunden, die in ordovizisch-silurischen epikontinentalen Meeresstraßen abgelagert wurden45.
Typischerweise kann ein niedriger TOC-Gehalt durch eine begrenzte Ablagerung von organischem Material in Umgebungen mit geringer Produktivität, ein geringes Konservierungspotenzial von organischem Material in der Wassersäule oder in den Sedimenten oder eine Verdünnung durch einen hohen silikatischen Fluss verursacht werden. In den modernen Ozeanen wird die Produktivität am häufigsten durch einen geringen Gehalt an essenziellen Nährstoffen begrenzt: Stickstoff, Phosphor, Silizium und Eisen46. Stickstoff, Phosphor und Eisen sind in Regionen mit offenem Ozean begrenzt, wo Auftrieb, Staubeintrag und Küstenabfluss keine ausreichenden Nährstoffmengen liefern. In modernen oligotrophen Gebieten ist das Verhältnis von bakterieller zu eukaryotischer Biomasse höher als in eutrophen oder mesotrophen Gebieten. Interessanterweise handelt es sich bei den Podillya 16PL-Aufschlussproben aus dem Redkino-Horizont von Moldawien um phosphorithaltige Schlammsteine, die ein deutlich niedrigeres Hopan/Steran-Verhältnis (8,1:11,5) aufweisen als die anderen Redkino-Proben in Tabelle 1 (obwohl sie immer noch deutlich höher sind als die im Salzbecken von Süd-Oman oder in organisch reichen phanerozoischen Sedimenten, die typischerweise in einen engen Bereich von 0.5-2,0 liegen), was möglicherweise darauf hindeutet, dass eine erhöhte Phosphatverfügbarkeit das lokal gefundene Verhältnis von Eukaryoten zu Bakterien günstig beeinflusst haben könnte (da die Stickstoffisotopensignaturen weitgehend unveränderlich sind, siehe nächster Abschnitt). Der Phosphorgehalt (P) sowie die Verhältnisse von P/Gesamt und P/Al sind in den Sedimenten des späten Ediacaran in Podillya, der Ukraine und Estland47 generell niedrig, mit Ausnahme dieser stratigraphischen Ebene, die durch Phosphoritknollen gekennzeichnet ist. Ähnlich niedrige P-Gehalte (~0,01 bis ~0,1 Gew.-%) in den siliklastischen Gesteinen von Kotlin und Redkino wurden aus einem Bohrkern vom nordöstlichen Rand der osteuropäischen Plattform11 gemeldet.
Die breite und flache Topographie in den epikontinentalen Meeren quer durch Baltika könnte die Phosphor- oder andere Nährstoffbeschränkungen in Randbereichen aufrechterhalten haben, und zwar aufgrund der authigenen Ausfällung von Phosphat mit Eisenmineralen in den oxischen Oberflächengewässern und der Sequestrierung einer Reihe von Elementen durch Schelfsedimente48. Wenn Phosphat ein limitierender Nährstoff war, hätte eine Erhöhung des bioverfügbaren Phosphors die lokale Primärproduktion gesteigert und günstigere Wachstumsbedingungen für größeres einzelliges Phytoplankton geschaffen49,50,51. In Bezug auf das moderne Ozeansystem ist bekannt, dass die marinen Picocyanobakterien, Prochlorococcus und Synechococcus, die Zellzahl und Biomasse des Phytoplanktons in den oligotrophen tropischen und subtropischen Ozeanen dominieren, einschließlich der phosphatbegrenzten oligotrophen Regionen des Oberflächenwassers52. Prochlorococcus und die heterotrophen SAR11 (Pelagibacter) gedeihen aufgrund einer Vielzahl von Anpassungen, darunter niedrige Energiekosten durch kleine Genome und niedrige Replikationsraten, ein größeres Verhältnis von Oberfläche zu Volumen durch kleinere Zellgrößen und zusätzliche Zellaufnahmefunktionen zur Maximierung der Nährstoffnutzung53. Viele Bakterien sind auch in der Lage, Nährstoffe in geringer Menge zu ersetzen, z. B. durch die Nutzung von Sulfolipiden anstelle von phosphorhaltigen Lipiden in einer Umgebung mit P-Mangel54 oder durch die Verwendung alternativer Substrate, z. B. durch die Beschaffung von Stickstoff aus atmosphärischem N2 über Stickstofffixierung55, um Nährstoffstress zu mildern. Eine neuere Studie über das Überleben von marinem Bakterioplankton in oligotrophen Umgebungen56 , in denen das verfügbare Phosphat begrenzt ist, deutet auf eine wichtige Rolle des Polyphosphat-Stoffwechsels in oligotrophen Meeresorganismen hin. Bei den ausgewählten Eukaryoten, die in oligotrophen Umgebungen konkurrieren, handelt es sich in der Regel um kleine Picoeukaryoten, die ihren Nährstoffbedarf durch Mixotrophie ergänzen können57. Das Verhältnis von Picoeukaryoten zu Cyanobakterien nimmt bei erhöhter Nährstoffzufuhr tendenziell zu49. Der mögliche Einfluss von Phosphor und anderen biolimitierenden Nährstoffen auf die Mäßigung der Primärproduktivität und die Struktur der Meeresgemeinschaften in epeirischen Meeren des Ediacaran muss weiter untersucht werden.
Ausnahmslos ist die Häufigkeit von C29-Steranen größer als die der entsprechenden C27- oder C28-Sterane für alle unsere Standorte (Tabelle 1). Ein Überwiegen von C29- gegenüber C27- und C28-Steranen deutet wahrscheinlich auf eine Dominanz von Grünalgen innerhalb der eukaryontischen Phytoplanktongemeinschaft hin44,58. Dieses Merkmal wurde in den meisten früheren Ediacaran-Biomarker-Studien beobachtet15,30,31,32,33,34,35,36,44. Bemerkenswert ist, dass die C30-Steran-Verteilung in mehreren Proben aus jedem Bohrkern im Kotlin-Horizont geringe, aber nachweisbare Mengen des Demosponge-Steran-Biomarkers 24-Isopropylcholestan (24-ipc) enthält59,60. Insgesamt wurden 24-ipc-Sterane in Gesteins- und Ölproben nachgewiesen, die bis ins Kryogenium (vor >635 Ma) im Salzbecken von Süd-Oman zurückreichen, und stellen die ältesten Lipid-Biomarker-Nachweise für Metazoen dar59,60. Die 24-ipc-Biomarker in unseren Proben waren entweder um eine Größenordnung weniger häufig (im Verhältnis zu den gesamten C27- zu C30-Steran-Verhältnissen lagen diese nur bei 0,06-0,61 %; Mittelwert = 0,22 %) im Vergleich zu Ediacaran-Gesteinen und -Ölen aus dem Süd-Oman (durchschnittlich 1,7 %59) oder lagen aufgrund ihrer vernachlässigbaren Häufigkeit bei der Mehrzahl der Proben unter der Nachweisgrenze.
Stickstoff- und organische Kohlenstoff-Isotopenverhältnisse
Stickstoff-Isotope können helfen, das relative Gleichgewicht im Stickstoffkreislauf zu erkennen und das Ausmaß zu bestimmen, in dem entweder die Stickstofffixierung oder die unvollständige Denitrifikation signifikante Wege waren, um die für marine Gemeinschaften verfügbare Nährstoffbilanz zu beeinflussen. Wenn diazotrophe Bakterien aufgrund eines Mangels an fixiertem Stickstoff in der Wassersäule molekularen Stickstoff fixieren, kann dies zu sedimentären Massenstickstoff-Isotopenwerten nahe 0‰61 führen. Dies steht im Gegensatz zu den positiven Stickstoffisotopensignaturen (im Bereich von +2 bis +10‰) mit einem Modus von +4 bis +6‰, die für die marinen Sedimente des Neoproterozoikums gefunden wurden, die unter Bedingungen abgelagert wurden, von denen man annimmt, dass sie nitratreich sind und Nitrat nur eine teilweise Denitrifikation erfahren hat62. Die Stickstoff-Isotopenwerte für alle unsere Proben mit Ausnahme eines Ausreißers (Tabelle 1) decken einen begrenzten positiven Bereich von +3,5 bis +6,5‰ ab, der sich mit dem Modus für spätneoproterozoische, an organischer Materie reiche Sedimentgesteine überschneidet62. In Verbindung mit Redox-Proxies, die darauf hindeuten, dass oxische Bedingungen in der Wassersäule vorherrschten, deuten unsere Daten darauf hin, dass Nitrat den gelösten anorganischen Stickstoffpool dominierte. Die Bandbreite der δ15N-Werte deutet auch darauf hin, dass die N2-Fixierung nicht die primäre Form der Stickstoffgewinnung für Primärproduzenten war. Vielmehr wurde der Stickstoffkreislauf wahrscheinlich von der Wiederverwertung in der Wassersäule dominiert, und der organische Stickstoff unterlag einer quantitativen Oxidation zu Nitrat, wie es auch in modernen, proximalen Meeresumgebungen der Fall ist. Die benthische Denitrifikation und die Vergrabung von organischem N wären die primären Senken für gelösten anorganischen Stickstoff gewesen, und die Bandbreite der δ15N-Werte deutet darauf hin, dass die unvollständige Denitrifikation in der Wassersäule, die in der Regel zu einer erheblichen 15N-Anreicherung führt, nur eine begrenzte Rolle spielt63. Die begrenzte Variabilität der δ15N-Werte spiegelt daher wahrscheinlich das δ15N von Nitrat wider, das auf die Plattform gelangt ist, mit einem geringen Einfluss der begrenzten Nitratreduktion und N2-Fixierung in der Wassersäule. Da es keine eindeutigen δ15N-Nachweise für die Stickstofffixierung gibt, gehen wir davon aus, dass Stickstoff nicht der primäre biolimitierende Nährstoff in den epikontinentalen Becken von Baltica war. Die Isolierung von flussnahen und äolischen Phosphorquellen und die oxischen Bedingungen in den weiten, flachmarinen epikontinentalen Becken könnten die Entfernung von authigenem Phosphorit und spurenmetallhaltigen Phasen gefördert haben, was zu einer Nährstofflimitierung führte, die das Wachstum und die Produktion eukaryotischer Zellen einschränkte.
Die Isotopenwerte des gesamten organischen Kohlenstoffs (CTOC) reichen von -23,0 bis -33,9‰, wobei die größten Unterschiede zwischen den verschiedenen Bohrkernstandorten beobachtet wurden. Die relative 13C-Anreicherung bei Isotopenverhältnissen über ca. -28‰ steht im Gegensatz zu Daten aus zeitgleichen Schichten, die in eutrophierten, offen-marinen Gebieten in Oman abgelagert wurden30,31. Die von uns angegebene Spanne stimmt jedoch im Allgemeinen mit der δ13CTOC-Spanne für andere Standorte in Baltica überein, die im selben Zeitraum abgelagert wurden11. Der Unterschied zwischen Baltica und Oman könnte die Tatsache unterstreichen, dass die δ13CTOC-Werte nicht ausschließlich eine einheitliche säkulare Veränderung im Kohlenstoffzyklus während dieses Zeitraums widerspiegeln31,64. Vielmehr könnte die δ13CTOC-Spanne von Baltica zum Teil die bakteriell dominierte mikrobielle Ökologie widerspiegeln, die durch die einzigartigen Lipid-Biomarker-Verhältnisse nahegelegt wird. Kleine Zellgrößen, ein hohes Verhältnis von Oberfläche zu Volumen und langsame Wachstumsraten unter oligotrophen Bedingungen können das Ausmaß der Fraktionierung während der Autotrophie (εp)65,66 erhöhen, was zu niedrigen δ13C-Werten führt.
Der Mechanismus für die stärker mit 13C angereicherten Isotopensignaturen innerhalb des für die Baltica-Sukzession im späten Ediacaran berichteten Bereichs11 ist weniger klar. Der mögliche Beitrag von detritalem, metamorph verändertem organischem Material zu präkambrischen Sedimentabfolgen mit niedrigem TOC-Gehalt ist ein möglicher Mechanismus für die Unterschiede zwischen den Standorten im δ13CTOC64. Der TOC-Gehalt unserer Proben ist zwar niedrig, liegt aber im Allgemeinen über 0,10 Gew.-%, so dass kein Zusammenhang zwischen TOC-Gehalt und δ13C-Werten erkennbar ist. Darüber hinaus enthalten unsere Proben thermisch unreifes organisches Material und Biomarker-Lipidmuster, die nicht mit einer hauptsächlich allochthonen Kohlenstoffquelle vereinbar sind. Stärker mit 13C angereicherte δ13CTOC-Signaturen können das Ergebnis alternativer Mechanismen der Kohlenstoffassimilation sein. Mechanismen der Kohlenstoffkonzentration oder der aktiven Bikarbonataufnahme durch Prokaryoten können zu kleineren Werten für εp und höheren δ13CTOC-Werten führen67,68. Die Entstehung der Ediacaran-Biota könnte die marinen Nahrungsnetze erheblich erweitert und neue Wege der mikrobiellen Heterotrophie stimuliert haben, einschließlich möglicher Beiträge aus komplexen Kohlenstoffkreisläufen innerhalb benthischer mikrobieller Matten. Unter zusätzlicher Berücksichtigung der potenziell wichtigen Rolle gelöster organischer Stoffe69 als Kohlenstoffquelle spiegelt die Bandbreite der δ13CTOC-Werte wahrscheinlich eine Reihe von biogeochemischen Kohlenstoffkreislaufprozessen wider, die für die sich entwickelnde Meeresumwelt des Ediakars einzigartig sein könnten und möglicherweise mit einem Phänomen zusammenhängen, für das wir keine repräsentativen modernen Analoga haben.
In den Gesteinen der frühen Trias in Südchina70 wurden alte Lipid-Biomarker und stabile Isotopen-Indikatoren für die benthische, mikrobielle Mattenproduktion in Verbindung mit einer Begrenzung des gebundenen Stickstoffs gefunden. Nach dem Massenaussterben am Ende des Perms kam es in Meishan zu einem starken Anstieg der Hopan/Steran-Verhältnisse (bis zu ca. 60), begleitet von einer starken Verschiebung der Norg-Isotopensignatur auf Werte von 0 bis -2‰, was auf bakterielle Diazotrophie hindeutet. Dies geht einher mit einem hohen Anteil an 2-Methylhopanen (2-Methylhopan-Index bis zu 33 %) und ausgeprägten Methylalkanen, die auf die Ausbreitung benthischer mikrobieller Matten hinweisen. Wir beobachten keinen ähnlichen Trend in der Stickstoffisotopensignatur oder in den Biomarker-Mustern, die offensichtlich auf einen signifikanten mikrobiellen Matteneintrag in unserem Ediacaran-Datensatz hinweisen würden, obwohl ein einzelner 15N-verarmter Wert in unseren Proben gefunden wurde (Tabelle 1). Daher ist das starke bakterielle Signal, das in unseren Baltica-Proben beobachtet wurde, wahrscheinlich nicht in erster Linie eine Signatur benthischer mikrobieller Matten.
Paläoumweltliche Ernährung der Ediacara-Biota vs. Demosponges Demosponges
Die offensichtlichen oligotrophen Bedingungen in den epikratonischen und kontinentalen Randbecken von Baltica, als es vom späten Ediacaran bis zum frühen Kambrium von den hohen zu den niedrigen Breiten driftete, könnten entweder mit der begrenzten Advektion von relativ nährstoffreichem Tiefenwasser oder mit der Verarmung an Nährstoffen in Verbindung stehen, die aus der Assimilation und dem Aufsaugen während des Transports und der Ablagerung über diese breiten, flachmarinen epikontinentalen Becken resultierte (Abb. 3). Diese Becken waren episodisch von den Ozeanen isoliert und entwickelten Hypersalinität (z. B. während der Redkino-Zeit) und brackische Bedingungen (z. B. während der Kotlin-Zeit71). Die langfristige tektonische Stabilität führte zu einer tiefliegenden Topographie des späten Ediacaran Baltica, die sehr anfällig für Überschwemmungen und eine ineffiziente Zufuhr von Phosphor aus der Verwitterung war. Im Gegensatz zu Baltica ergaben Biomarker-Untersuchungen der Schichten der Huqf-Supergruppe in Oman ein eutrophes Ökosystem, das reich an Mikroalgen war30,31,59, aber keine Ediacara-Biota aufwies, selbst nicht in den ausgedehnten Aufschlüssen in den Oman-Bergen und der Huqf-Region, in den Bereichen des inneren und äußeren Schelfs3. Die Paläogeographie des Süd-Oman-Salzbeckens wurde für das späte Neoproterozoikum auf ~13° vom Äquator in der südlichen Hemisphäre rekonstruiert, was weitgehend der Baltica72-Paläolatitude entspricht.
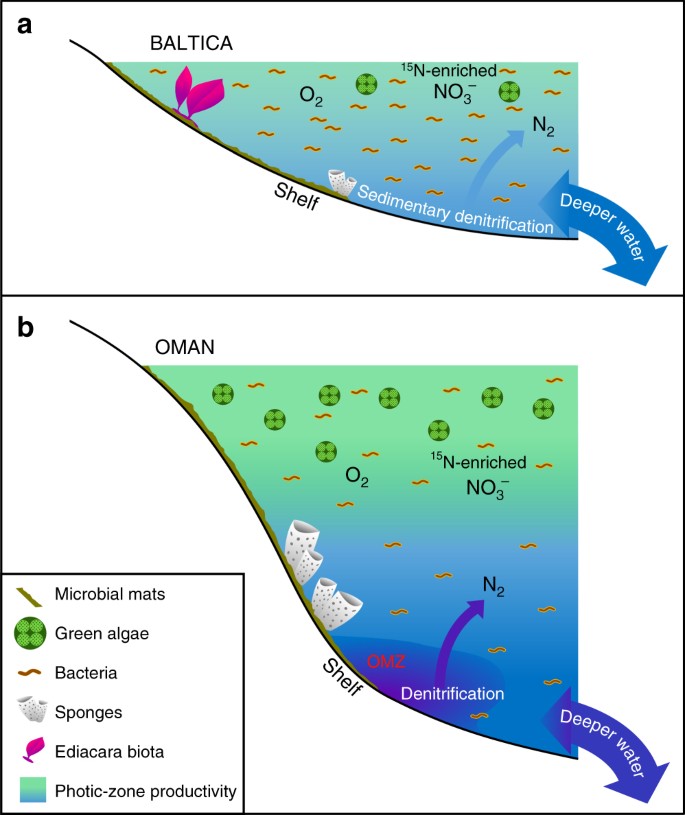
Hauptunterschiede zwischen wenig produktiven und produktiven Ediacaran-Meeresumgebungen. Schematische Diagramme werden gezeigt für a ausgedehnte oligotrophe und flachmarine epikontinentale Beckenränder von Baltica, die oft von bakterieller Produktivität dominiert wurden, wo die Ediacara-Weichkörperfauna florierte und wo Denitrifikation und Anammox wahrscheinlich auf die Sedimente beschränkt waren; und b eutrophe und tiefer gelegene Schelfgebiete des Süd-Oman-Salzbeckens, wo Grünalgen als Primärproduzenten gediehen und Demosponges reichlich vorhanden waren, aber keine Ediacara-Weichkörperfauna auftrat (wobei Ediacara-Biota-Fossilien auch in entsprechenden Ediacaran-Aufschlüssen im nördlichen Oman fehlten). An produktiven Kontinentalrändern fanden Denitrifikation und Anammox wahrscheinlich sowohl in der Wassersäule als auch in den Sedimenten statt
Es gibt keine Beweise für anhaltende und ausgedehnte Anoxie an und um die flachen Kontinentalränder von Baltica während des späten Ediacaran11, und wir vermuten, dass die hier beschriebenen oligotrophen Bedingungen durch ineffiziente terrestrische und Tiefwasser-Nährstoffflüsse in diese Gebiete verursacht wurden, die im Großen und Ganzen denen in oligotrophen Umgebungen in modernen Ozeansystemen ähneln. Eine geochemische Spurenelement-Untersuchung des Utkina Zavod und angrenzender Bohrkerne aus der Gegend von St. Petersburg legt nahe, dass unsere Proben unter oxischen Bedingungen abgelagert wurden73. Diese Interpretation wird unabhängig von unseren Daten durch die erweiterten Hopanverteilungen (C31-C35) gestützt, deren Häufigkeit oberhalb von C31-Verbindungen mit zunehmender Kohlenstoffzahl stark abnimmt, was für den Seitenkettenabbau von Bakteriohopanpolyolen während der Diagenese unter oxischen Bedingungen charakteristisch ist29, und auch durch niedrige Wasserstoffindizes, wie sie durch die Rock-Eval-Pyrolyse gemessen wurden (alle unter 230 mg/g TOC und viele unter 100 mg/g TOC), für diese unreifen Proben aufgrund der Bildung von rekalzitantem Kerogen durch oxidativen Abbau und Rekondensation von lipidarmer primärer Biomasse in lokal oxischen Umgebungen (Tabelle 1). Die mehrzelligen Organismen des Ediacaran, die in diesen flachmarinen Umgebungen lebten, müssen über ausreichend organische Substrate für die Heterotrophie verfügt haben, um ihren Nahrungsbedarf zu decken, sowie über genügend Sauerstoff und andere Nährstoffe, um ihren Stoffwechsel aufrechtzuerhalten. Epikontinentale Becken von Baltica waren wahrscheinlich dauerhaft sauerstoffhaltiger als die hochproduktiven Gebiete, die Sauerstoffminimumzonen säumten, wie z. B. auf dem mittleren bis äußeren Schelf des Salzbeckens von Süd-Oman, wo die Atmung der reichlich vorhandenen planktonischen Biomasse einen niedrigeren gelösten Sauerstoff unterhalb der photischen Zone aufrechterhalten hätte.
Ediacara-Biota und andere mehrzellige Organismen, die vor der Küste von Baltica leben, mussten mit wechselnden Nahrungsquellen (bakteriell vs. eukaryotisch) zurechtkommen, da die Nährstoffflüsse im Laufe der Zeit variierten, einschließlich kleiner Zellen und organischem Detritus in epikontinentalen Becken, wo Bakterien die dominierenden Primärproduzenten waren (Abb. 3). Moderne Korallenriff-Gemeinschaften überleben in tropischen oligotrophen Umgebungen aufgrund des effizienten Recyclings von Nährstoffen, einschließlich der Erzeugung eines Flusses von gelösten organischen Stoffen (DOM), der zur Aufrechterhaltung der Heterotrophie der Fauna innerhalb des Riff-Ökosystems beiträgt74. Die Etablierung einer marinen trophischen Struktur mit eukaryotischen mehrzelligen Organismen, die sich von organischem Detritus ernähren, musste nach der globalen Ausbreitung von Eukaryoten in verschiedene Meeresumgebungen erfolgen, die im Tonian-Cryogenian-Intervall (ca. 800-635 Ma) stattfand, wie aus Biomarker-Aufzeichnungen ersichtlich ist15. Es ist zu erwarten, dass wir während des späten Ediacarans von Ort zu Ort signifikante Unterschiede in der Struktur der marinen Gemeinschaften und der Trophie feststellen können, wobei die lokale Nährstoffbilanz eukaryotenreiche oder eukaryotenarme mikrobielle Gemeinschaften begünstigt und die fortschreitende Ausbreitung mehrzelliger Organismen eine weitere Dimension der Komplexität auf der Ebene der Organismen und Gemeinschaften hinzufügt. Erhebliche regionale Unterschiede in der Phosphat- und anderen Nährstoffverfügbarkeit in Schelfumgebungen sind auch eine erwartete Folge einer heterogenen globalen marinen Redoxstruktur im späten Ediacaran, vor der Sauerstoffanreicherung der Tiefsee38.
Die Dominanz der bakteriellen Produktivität des Picoplanktons und der damit verbundenen Abbauprodukte gelöster organischer Stoffe (DOM) in den epikontinentalen Becken der Baltica könnte ein mikrobielles Kreislaufökosystem parallel zu der konventionellen trophischen Struktur aufrechterhalten haben, die auf größeren planktischen Produzenten basiert50. Dies könnte verschiedene Formen der Heterotrophie begünstigt haben, darunter Suspensionsfütterung und möglicherweise Osmotrophie als praktikable Ernährungsstrategie für einige Rangeomorphe, Schwämme und andere multizelluläre Organismen des späten Ediacaran18 in Verbindung mit der aufkommenden aktiven (beweglichen) Heterotrophie19,20. Nährstoffbegrenzte aquatische Systeme werden in der Regel von kleinem einzelligem Phytoplankton und heterotrophem Plankton dominiert, wobei Bakterien die Eukaryoten verdrängen und in oligotrophen marinen Umgebungen des modernen Ozeans eine geringe Nettobiomasse aufrechterhalten49,51. DOM ist eine wichtige Quelle organischer Nährstoffe und steuert häufig die Produktivität und die Nettobiomasse in modernen oligotrophen tropischen Meeren, aber auch die Ko-Limitierung von Stickstoff und Phosphor75. In ähnlicher Weise war DOM wahrscheinlich auch ein wichtiges Substrat für die Erhaltung heterotropher Bakterien und eines mikrobiellen Kreislaufs in alten oligotrophen Umgebungen. Benthische mikrobielle Matten könnten ein Bestandteil dieses bakteriell dominierten Nahrungsnetzes gewesen sein, und Mattenböden haben die Erhaltung der Ediacara-Biota in mariner Umgebung gefördert7. Während für eine Teilmenge unserer Proben mäßige Werte des 2-Methylhopan-Index gefunden wurden (4-10 %, Tabelle 1), deuten die sehr geringen Mengen an Methylalkanen im Vergleich zu n-Alkanen (Abb. 1) und das nur sporadische Vorkommen von Carotinoiden darauf hin, dass mikrobielle Matten die Primärproduktivität nicht dominierten76 und dass planktonische Bakterien und dementsprechend ihre DOM-Abbauprodukte florierten. Die allgemein niedrigen Wasserstoff-Index-Werte (Tabelle 1; insbesondere für die Redkino-Proben) stehen trotz der geringen thermischen Reife der Schichten und Biomarker-Assemblagen ebenfalls im Einklang mit lipidarmen organischen Einträgen in überwiegend oxischen, flachmarinen Randbereichen. Das Vorkommen von mikrobiellen Matten, das nur anhand der sedimentologischen Textur gemessen wird, sagt nichts über das Verhältnis von Eukaryoten (Mikroalgen) zu Bakterien aus und kann auch nicht den relativen Beitrag des mikrobiellen Planktons zur Gesamtprimärproduktivität und zum Unterhalt der Nahrungsnetze einschränken. Mikrobielle Mattengemeinschaften, die oft reichlich Eukaryoten enthalten, und die Karbonate der Ara-Gruppe aus Süd-Oman mit ausgeprägten thrombolitischen und faltigen Laminit-Fazies sind ein gutes Beispiel für spätneoproterozoische Umgebungen mit einem bedeutenden Beitrag mikrobieller Matten, die aufgrund eines großen Beitrags von Mikroalgen reichlich Steran-Signale liefern30,59.
Die extrem niedrige Konzentration von 24-ipc-Biomarkern deutet darauf hin, dass Demosponges zwar manchmal vorhanden waren, aber in diesen Umgebungen nur wenig. Dies könnte darauf hinweisen, dass kleinere filtrierende Tiere, die mit der Ediacara-Biota konkurrieren, in diesen Umgebungen mit geringer Produktivität weniger Chancen haben, dass die Demosponges nicht ausreichend widerstandsfähig gegen die energiereicheren Bedingungen im flachen Meer oberhalb der Schönwetter-Wellenbasis sind oder dass die Schwämme besser an die sauerstoffarmen Bedingungen angepasst sind, die in eutrophen Umgebungen dynamisch unterhalb der photischen Zone aufrechterhalten werden (Abb. 3). Insgesamt wurde 24-ipc in den meisten Proben des Redkino-Horizonts, die im Allgemeinen auch die höchsten H/St-Verhältnisse in unserem Probensatz aufweisen, nicht nachgewiesen, was möglicherweise auf eine ökologische Veränderung während der Ablagerungszeit des Kotlin-Horizonts hinweist. Alternativ ist es auch plausibel, dass Schwämme diese Gebiete bewohnten, aber diese diagnostischen Steroid-Biomarker nicht im Überfluss produzierten, obwohl dies weniger wahrscheinlich erscheint, da 24-ipc-Steran zu den am häufigsten nachgewiesenen C30-Steranverbindungen in Ediacaran-Schichten und -Ölen gehört30,3133,59,60 und auch in einer Untergruppe unserer Proben nachweisbar ist (Tabelle 1).
Die anhaltenden oligotrophen Meeresumgebungen, auf die unsere Daten hindeuten, stellen zwar lokal begrenzte Bedingungen in den Ediacaran-Ozeanen dar, waren aber für präkambrische flachmarine Meeresstraßen wahrscheinlich nicht ungewöhnlich (Abb. 3). Die überwiegende Mehrheit der bisherigen Ediacaran-Biomarker-Studien wurde an organisch reichen Sedimentgesteinen durchgeführt, die in eutrophem Milieu abgelagert wurden, sowie an deren Erdölprodukten, die im Allgemeinen Biomarker-Assemblagen ergeben, die auf einen signifikanten Beitrag von Mikroalgen schließen lassen44. Unsere Ergebnisse machen deutlich, wie wichtig es ist, eine größere Vielfalt an Ablagerungsumgebungen zu untersuchen, einschließlich organisch-magerer Schichten mit angemessener thermischer Reife und unterschiedlicher Lithologie15,3034,59, um ein genaueres Bild vom Ausmaß der Heterogenität der Meereschemie und -ökologie von Ort zu Ort zu erhalten. Trotz der fortschreitenden Belüftung des Ozeans und der zunehmenden chemischen Verwitterung und Nährstoffzufuhr während des Auseinanderbrechens von Rodinia und während der gesamten Ediacaran-Periode77,78 sorgte die Heterogenität des Ozeans für die Aufrechterhaltung einer Vielzahl von chemischen Bedingungen im Meer, einschließlich nährstoffarmer, aber bewohnbarer Umgebungen, die die Anpassung, den Wettbewerb und die Evolution von Metazoen innerhalb des globalen Ozeansystems förderten (Abb. 3). Während eutrophe marine Schelfe Demosponges beherbergten, aber oft nicht die Ediacara-Biota, wobei das Süd-Oman-Salzbecken (und die entsprechenden dicken Ediacaran-Aufschlüsse weiter nördlich in Oman) ein prominentes Beispiel ist; kontraintuitiv wurden flache, oxische und weniger produktive epikontinentale Meeresstraßen von der Ediacara-Biota bevorzugt von Demosponges besiedelt, obwohl DOM und anderer organischer Detritus lokal für die Ernährung verfügbar war. Ob Stoffwechselbedürfnisse oder Selektionsdruck aus der Umwelt die mehrzelligen Biota des Ediacaras auf diese Umgebungen beschränkt haben, bleibt ungewiss; unsere Studie unterstreicht jedoch, dass oligotrophe Umgebungen des Ediacaras eine potenziell entscheidende Rolle bei der Evolution makroskopischer mehrzelliger Organismen und der Ökologie mariner Gemeinschaften gespielt haben.