La madurez térmica excepcionalmente baja de los estratos
La madurez térmica es el factor más importante que influye en la conservación de los antiguos biomarcadores sedimentarios, ya que las moléculas orgánicas son susceptibles de sufrir alteraciones estructurales y estereoquímicas durante el progresivo enterramiento y con el aumento del estrés térmico29. Las rocas precámbricas con una madurez térmica adecuada (madurez de la ventana de petróleo o inferior) son requisitos previos para preservar conjuntos de lípidos biomarcadores robustos y primarios. Las múltiples proporciones de estereoisómeros de hopano y esterano, junto con la evidencia independiente de los bajos valores de Tmáx (en su mayoría dentro de un rango de 417-433 °C, con una media de 426 °C) de la pirólisis Rock-Eval, indican que las rocas de este estudio representan, con mucho, las rocas ediacaranas más inmaduras térmicamente analizadas hasta la fecha utilizando los métodos geoquímicos orgánicos más avanzados (Tabla 1). Aunque nuestras rocas han sufrido una diagénesis sedimentaria, no han pasado significativamente a la ventana de petróleo, aparte de las muestras ligeramente más maduras del afloramiento 16PL de la cuenca de Podillya, en Moldavia, que siguen siendo adecuadas para el análisis (madurez de la ventana de petróleo de temprana a media y sin signos evidentes de contaminantes orgánicos). Es probable que la mayor parte del conjunto no haya estado expuesta a temperaturas de enterramiento superiores a 50 °C, por lo que nuestras muestras son muy inmaduras. Otras líneas de evidencia molecular, que apoyan la baja madurez térmica y la singenicidad, incluyen (i) un predominio de los alcanos policíclicos biomarcadores sobre los n-alcanos en los extractos de roca (Fig. 2), (ii) la supervivencia de cantidades detectables de 17β,21β(H)-hopanos resolubles a partir de los hopanos más abundantes que poseen configuraciones estables 17β,21α(H)- y 17α,21β(H)-estereoquímicas (Tabla 1, Figs. Suplementarias. 1 y 2), (iii) una preferencia discernible entre los n-alcanos en el rango de C22 a C27 (Fig. 2), ya que los n-alcanos muestran una preferencia por el número de carbono sólo antes de la catagénesis38, y (iv) la generación de biomarcadores de hopano y esterano térmicamente inmaduros a partir de la fase de kerógeno (insoluble) utilizando hidropirólisis catalítica (Fig. 2 suplementaria). La baja madurez térmica de la materia orgánica sedimentaria en nuestras muestras es consistente con los indicadores de alteración sedimentaria previamente publicados, incluyendo los índices de alteración del conodonto y del acritarco, y los estudios previos de pirólisis y biomarcadores Rock-Eval de las rocas sedimentarias ediacaranas y paleozoicas del Báltico36,39,40, así como la mineralogía de las arcillas41. Críticamente, en entornos donde la madurez térmica es tan baja, los mecanismos para la alteración significativa de las señales primarias δ15N y δ13C están en gran medida ausentes42.
Tabla 1 Seleccione las proporciones de biomarcadores lipídicos para la madurez térmica, la biota de origen y las evaluaciones ambientales de depósito
Fig. 2
Distribución de hidrocarburos alifáticos extraíbles para una muestra representativa. a Cromatograma de iones totales (TIC) para hidrocarburos alifáticos extraíbles para Lugovoe #13-73 m del Horizonte Redkino. La serie de n-alcanos, el pristano (Pr), el fitano (Ph) y los hopanos C27-C34 (indicados por su número total de carbono y su estereoquímica en C-17, C-21 y C-22, por ejemplo, C31αβR) están marcados; ** indica los hopenos C29 y C30. Obsérvese el predominio de la abundancia de hopenos sobre otras clases de compuestos alcanos. b El cromatograma parcial de iones 85-Da muestra el predominio de la serie de n-alcanos, exhibiendo una preferencia discernible de números de carbono impares sobre pares en el rango C22-C27, sobre los metilalcanos
Patrones de ensamblaje de biomarcadores de lípidos
Una relación de abundancia de los hopanos principales (C27-C35) con respecto a los esteranos principales (C27-C29) se utiliza a menudo para evaluar el equilibrio de los aportes de organismos fuente bacterianos frente a los eucariotas al antiguo ecosistema acuático. Los hopanos son fósiles moleculares derivados de los hopanoides, que son lípidos de la membrana celular sintetizados por una amplia variedad de grupos bacterianos. Del mismo modo, los esteranos se derivan de precursores de esteroles, que son producidos casi exclusivamente por eucariotas43. Los ratios H/St de nuestras muestras cubren un rango sorprendentemente amplio de valores, desde 1,6 hasta 119,2 (Tabla 1). Para contextualizar, las relaciones H/St de rocas y aceites neoproterozoicos ricos en materia orgánica se sitúan normalmente en un estrecho rango de 0,5 a 2,030,31,32,33. Mientras hay una diferencia clara en los valores del Horizonte Kotlin más joven (H/St promedio de 8.9) y el Horizonte Redkino más viejo (H/St promedio de 42.9), los valores para la mayoría de las muestras son inusualmente altos y sugieren contribuciones anormalmente elevadas de bacterias.
Para el Neoproterozoico tardío, las algas eucariotas eran un componente ecológicamente significativo y productores principales en muchos ecosistemas marinos15,34,44. La discrepancia entre la contribución globalmente importante de las algas eucariotas en el Neoproterozoico tardío y, sin embargo, los bajos niveles de biomarcadores de esterano en estas muestras indica que debe haber algún determinante local en la abundancia de eucariotas. La explicación más parsimoniosa para las elevadísimas relaciones hopano/esterano junto con los bajos contenidos de carbono orgánico total (COT) y los bajos índices de hidrógeno (IH) encontrados en nuestras muestras (Tabla 1) es que estos estratos se depositaron en entornos oligotróficos (es decir, fuertemente limitados por los nutrientes), en los que las bacterias superaron a las algas. Mientras que los análogos modernos para los antiguos mares epicratónicos desarrollados durante el alto nivel del mar son difíciles de encontrar, se han encontrado previamente observaciones paralelas de proporciones más altas de hopano/esterano (por hasta un orden de magnitud) para las rocas sedimentarias magras orgánicas frente a las ricas en materia orgánica depositadas en las vías marítimas epicontinentales del Ordovícico-Silúrico45.
Típicamente, el bajo contenido de COT puede ser causado por la deposición limitada de materia orgánica en entornos de baja productividad, el bajo potencial de preservación de la materia orgánica en la columna de agua o en los sedimentos, o la dilución con un alto flujo siliciclástico. En los océanos modernos, la productividad suele estar limitada por los bajos niveles de los nutrientes esenciales: nitrógeno, fósforo, silicio y hierro46. El nitrógeno, el fósforo y el hierro están limitados en las regiones de mar abierto donde el afloramiento, la entrada de polvo y la escorrentía costera no suministran cantidades suficientes de nutrientes. En los entornos oligotróficos modernos, la relación entre la biomasa bacteriana y la eucariótica es mayor que en los entornos eutróficos o mesotróficos. Curiosamente, las muestras del afloramiento Podillya 16PL del horizonte Redkino de Moldavia son fangolitas que contienen fosforita y arrojan proporciones de hopano/esterano significativamente más bajas (8,1:11,5) que las otras muestras de Redkino de la Tabla 1 (aunque siguen siendo significativamente más altas que las encontradas en la cuenca salina del sur de Omán o en los sedimentos fanerozoicos ricos en materia orgánica, que suelen estar dentro de un estrecho rango de 0.5-2,0), lo que posiblemente sugiere que el aumento de la disponibilidad de fosfato podría haber influido favorablemente en la relación eucariota-bacteria encontrada localmente (dado que las firmas isotópicas de nitrógeno son en gran medida invariables, véase la siguiente sección). Los contenidos de fósforo (P), así como las relaciones P/Fetotal y P/Al, son por lo demás generalmente bajos para los sedimentos ediacaranos tardíos de Podillya, Ucrania y Estonia47; con la excepción de este nivel estratigráfico marcado con nódulos de fosforita. Del mismo modo, los niveles bajos (~0,01 a ~0,1 wt%) de P en las rocas siliciclásticas de Kotlin y Redkino fueron reportados desde un núcleo de perforación del margen noreste de la Plataforma de Europa Oriental11.
La amplia y poco profunda topografía en los mares epicontinentales a través del Báltico podría haber sostenido limitaciones de fósforo u otros nutrientes en entornos marginales, debido a la precipitación autógena de fosfato con minerales de hierro en las aguas superficiales óxicas y el secuestro de una serie de elementos por los sedimentos de la plataforma48. Si el fosfato fuera un nutriente limitante, el aumento del fósforo biodisponible habría mejorado la producción primaria local y proporcionado condiciones de crecimiento más favorables para el fitoplancton unicelular de mayor tamaño49,50,51. Con respecto al sistema oceánico moderno, se reconoce que las picocyanobacterias marinas, Prochlorococcus y Synechococcus, dominan los recuentos celulares y la biomasa del fitoplancton en los entornos oligotróficos de los océanos tropicales y subtropicales, incluidas las regiones oligotróficas limitadas por el fosfato de las aguas superficiales del océano52. El Prochlorococcus y el heterótrofo SAR11 (Pelagibacter) prosperan gracias a una serie de adaptaciones, entre las que se incluyen un bajo coste energético en virtud de sus pequeños genomas y bajas tasas de replicación, una mayor relación superficie-volumen gracias a un menor tamaño de las células y funciones adicionales de captación celular para maximizar la utilización de los nutrientes53. Muchas bacterias también son capaces de sustituir los nutrientes de baja abundancia, por ejemplo, utilizando sulfolípidos en lugar de lípidos que contienen fósforo en entornos con deficiencia de P54, o mediante el uso de sustratos alternativos, por ejemplo, obteniendo nitrógeno del N2 atmosférico a través de la fijación de nitrógeno55, para aliviar el estrés de nutrientes. Un estudio reciente sobre la supervivencia del bacterioplancton marino en entornos oligotróficos56, donde el fosfato disponible es limitado, sugiere un importante papel del metabolismo del polifosfato en los oligótrofos marinos. Los eucariotas selectos que compiten en entornos oligotróficos son típicamente pequeños picoeucariotas, que pueden complementar sus necesidades nutricionales a través de la mixotrofia57. La proporción de biomasa entre picoeucariotas y cianobacterias tiende a aumentar bajo un mayor suministro de nutrientes49. La posible influencia del fósforo, y de otros nutrientes biolimitadores, en la moderación de la productividad primaria y de la estructura de las comunidades marinas en las vías marítimas epeiricas ediacaranas requiere una investigación más profunda.
Sin excepción, la abundancia de esteranos C29 es mayor que la de los esteranos C27 o C28 correspondientes para todas nuestras localizaciones (Tabla 1). El predominio de los esteranos C29 sobre los C27 y C28 indica probablemente un predominio de las algas verdes dentro de la comunidad de fitoplancton eucariota44,58. Esta característica ha sido observada en la mayoría de los estudios anteriores de biomarcadores ediacaranos15,30,31,32,33,34,35,36,44. En particular, la distribución del esterano C30 en varias muestras de cada núcleo de perforación en el horizonte Kotlin contiene cantidades bajas, pero detectables, del biomarcador de esterano de la demosponja conocido como 24-isopropilcolestano (24-ipc)59,60. En total, se ha informado de la presencia de esteranos de 24-ipc en muestras de roca y petróleo que se remontan al Criogénico (hace >635 Ma) en la cuenca salina del sur de Omán y representan la evidencia más antigua de biomarcadores lipídicos para los metazoos59,60. Los biomarcadores de 24-ipc en nuestras muestras eran, o bien de un orden de magnitud inferior en abundancia (en relación con las proporciones totales de esterano C27 a C30, éstas eran sólo de 0,06 a 0,61%; media = 0,22%) en comparación con las rocas y aceites ediacaranos del sur de Omán (1,7% de media59), o bien estaban por debajo de los límites de detección debido a una abundancia insignificante para la mayoría de las muestras.
Las relaciones isotópicas del nitrógeno y del carbono orgánico
Los isótopos del nitrógeno pueden ayudar a discernir el equilibrio relativo en el ciclo del nitrógeno, y el grado en que la fijación del nitrógeno o la desnitrificación incompleta eran vías importantes para influir en el equilibrio de los nutrientes disponibles para las comunidades marinas. Cuando las bacterias diazótrofas fijan el nitrógeno molecular debido a la falta de nitrógeno fijado en la columna de agua, esto puede dar lugar a valores sedimentarios de isótopos de nitrógeno en masa cercanos a 0‰61. Esto contrasta con las firmas isotópicas de nitrógeno positivas (en el rango de +2 a +10‰) con un modo de +4 a +6‰ encontradas para los sedimentos marinos del Neoproterozoico depositados bajo lo que se piensa que son condiciones de nitrato-repleto donde el nitrato sólo ha sufrido una desnitrificación parcial62. Los valores de isótopos de nitrógeno de todas nuestras muestras, salvo un valor atípico (Tabla 1), cubren un rango positivo limitado de +3,5 a +6,5‰, que coincide con el modo de las rocas sedimentarias ricas en materia orgánica del Neoproterozoico tardío62. Limitados por los proxies redox que sugieren que las condiciones de la columna de agua eran óxicas, nuestros datos implican que el nitrato dominaba la reserva de nitrógeno inorgánico disuelto. El rango de valores δ15N también sugiere que la fijación de N2 no era el modo principal de adquisición de nitrógeno para los productores primarios. Más bien, el ciclo del nitrógeno estaba probablemente dominado por el reciclaje de la columna de agua, y el N orgánico estaba sujeto a la oxidación cuantitativa a nitrato como lo está en los entornos marinos modernos y proximales. La desnitrificación bentónica y el enterramiento de N orgánico habrían sido los principales sumideros de nitrógeno inorgánico disuelto, y el rango de valores de δ15N sugiere sólo un papel limitado para la desnitrificación incompleta de la columna de agua, que típicamente resulta en un enriquecimiento significativo de 15N63. La limitada variabilidad de los valores de δ15N refleja, por tanto, el δ15N del nitrato que entra en la plataforma, con una influencia menor de la reducción del nitrato en la columna de agua y la fijación del N2. Sin una fuerte evidencia de δ15N para la fijación de nitrógeno, nuestra hipótesis es que el nitrógeno no fue el principal nutriente biolimitante en las cuencas epicontinentales del Báltico. El aislamiento de las fuentes fluviales y eólicas de fósforo, y las condiciones óxicas en las amplias cuencas epicontinentales marinas poco profundas pueden haber potenciado la eliminación de la fosforita autógena y de las fases portadoras de metales traza, dando lugar a una limitación de nutrientes que restringió el crecimiento y la producción de las células eucariotas.
Los valores isotópicos del carbono orgánico total (CTOC) oscilan entre -23,0 y -33,9‰, observándose las mayores diferencias entre las distintas localizaciones de los sondeos. El enriquecimiento relativo en 13C de las relaciones isotópicas superiores a aproximadamente -28‰ contrasta con los datos de estratos contemporáneos depositados en los entornos eutróficos y marinos abiertos de Omán30,31. Sin embargo, el rango que reportamos es generalmente consistente con el rango de δ13CTOC para otras localidades del Báltico depositadas durante el mismo período de tiempo11. La diferencia entre el Báltico y Omán podría resaltar el hecho de que los valores de δ13CTOC no reflejan exclusivamente un cambio secular uniforme en el ciclo del carbono durante este intervalo de tiempo31,64. Más bien, el rango de δ13CTOC del Báltico puede reflejar, en parte, la ecología microbiana dominada por bacterias que sugieren las relaciones únicas de los biomarcadores lipídicos. El tamaño pequeño de la célula, las altas relaciones superficie-volumen, y las tasas de crecimiento lentas bajo condiciones oligotróficas pueden aumentar la magnitud del fraccionamiento durante la autotrofia (εp)65,66, dando lugar a valores bajos de δ13C.
El mecanismo para las firmas isotópicas más enriquecidas en 13C dentro del rango reportado11 para la sucesión tardía del Ediacaran Baltica está menos claro. La contribución potencial de la materia orgánica detrital, metamórficamente alterada a las sucesiones sedimentarias precámbricas de bajo COT es un mecanismo posible para las disparidades entre sitios en δ13CTOC64. Sin embargo, el contenido de COT de nuestras muestras, aunque bajo, es generalmente superior al 0,10 % en peso, lo que no revela ninguna relación entre el contenido de COT y los valores de δ13C. Además, nuestras muestras contienen materia orgánica térmicamente inmadura y patrones de lípidos biomarcadores que son inconsistentes con una fuente de carbono principalmente alóctona. Las firmas de δ13CTOC más enriquecidas en 13C pueden ser el resultado de mecanismos alternativos de asimilación de carbono. Los mecanismos de concentración de carbono o de captación activa de bicarbonato por los procariotas pueden dar lugar a valores menores de εp y a valores más elevados de δ13CTOC67,68. La aparición de la biota ediacarana puede haber ampliado considerablemente las redes alimentarias marinas y haber estimulado nuevas vías de heterotrofia microbiana, incluidas las posibles contribuciones del complejo ciclo del carbono dentro de los tapetes microbianos bentónicos. Con la consideración adicional del papel potencialmente importante de la materia orgánica disuelta69 como fuente de carbono, la amplitud de los valores de δ13CTOC refleja probablemente una gama de procesos biogeoquímicos del ciclo del carbono que podrían ser únicos al medio ambiente marino ediacarano en evolución y pueden estar relacionados con un fenómeno para el cual no tenemos análogos modernos representativos.
Se han encontrado antiguos conjuntos de biomarcadores lipídicos e indicadores de isótopos estables para la producción de esteras microbianas bentónicas en tándem con la limitación de nitrógeno fijo en las rocas del Triásico temprano del Sur de China70. Tras la extinción masiva de finales del Pérmico en Meishan, se produjo un gran pico en las relaciones hopano/esterano (hasta aproximadamente 60) acompañado de un fuerte cambio en la firma isotópica de Norg a valores de 0 a -2‰, consistente con la diazotrofia bacteriana. Esto se asocia con una alta señal de 2-metilhopanos (índice de 2-metilhopano de hasta el 33%) y metilalcanos distintivos que apuntan a la proliferación de tapetes microbianos bentónicos. No observamos una tendencia similar en la firma de isótopos de nitrógeno o en los patrones de biomarcadores que obviamente apuntarían a una entrada significativa de tapetes microbianos dentro de nuestro conjunto de datos ediacaranos, aunque se encontró un valor singular de 15N agotado en nuestras muestras (Tabla 1). Por lo tanto, es probable que la fuerte señal bacteriana observada en nuestras muestras de la Báltica no sea predominantemente una firma de tapetes microbianos bentónicos.
Sostenimiento paleoambiental de la biota de Ediacara frente a las Las condiciones oligotróficas aparentes a través de las cuencas epicratónicas y de los márgenes continentales del Báltico, a medida que se desplazaba de las latitudes altas a las bajas desde el Ediacaran tardío hasta el Cámbrico temprano, podrían estar asociadas a una advección limitada de aguas profundas relativamente ricas en nutrientes o a un agotamiento de los mismos, como resultado de la asimilación y la depuración durante el transporte y la deposición a través de estas amplias cuencas epicontinentales de aguas poco profundas (Fig. 3). Estas cuencas fueron aisladas episódicamente de los océanos y desarrollaron hipersalinidad (por ejemplo, durante la época de Redkino) y condiciones salobres (por ejemplo, durante la época de Kotlin71). La estabilidad tectónica a largo plazo dio lugar a una topografía de bajo relieve de la Báltica ediacarana tardía, muy susceptible a las inundaciones y al suministro ineficaz de fósforo derivado de la meteorización. En contraste con el Báltico, los estudios de biomarcadores de los estratos del Supergrupo Huqf en Omán revelaron un ecosistema eutrófico, rico en microalgas30,31,59, pero carente de biota Ediacara, incluso en los extensos afloramientos de las Montañas de Omán y de la región del Huqf, en los entornos de la plataforma interior a la exterior3. La paleogeografía de la cuenca salina del sur de Omán ha sido reconstruida para el Neoproterozoico tardío a ~13° del ecuador en el hemisferio sur, ampliamente similar a la paleolatitud del Báltico72.
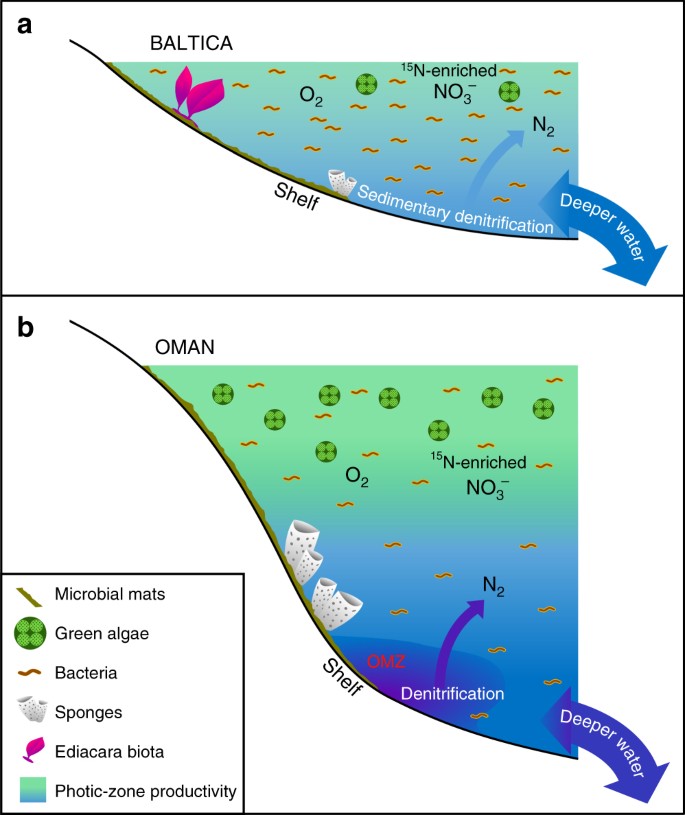
Fig. 3
Diferencias importantes en los medios marinos ediacaranos poco productivos frente a los productivos. Los diagramas esquemáticos se muestran para a extensos márgenes de cuencas epicontinentales oligotróficas y poco profundas del Báltico, a menudo dominados por la productividad bacteriana, donde floreció la fauna de cuerpo blando de Ediacara y donde la desnitrificación y el anammox probablemente estaban restringidos a los sedimentos; y b los entornos eutróficos y de plataforma marina más profunda de la cuenca salina del sur de Omán, donde las algas verdes prosperaban como productoras primarias y las demosponjas eran abundantes, pero la fauna de cuerpos blandos de Ediacara no era prominente (con fósiles de biota de Ediacara también ausentes en los afloramientos ediacaranos correlativos en el norte de Omán). En los márgenes continentales productivos, la desnitrificación y el anammox se produjeron probablemente tanto en la columna de agua como en los sedimentos
No hay pruebas de una anoxia persistente y extensa en los márgenes continentales poco profundos del Báltico y sus alrededores durante el Ediacaran tardío11, y sugerimos que las condiciones oligotróficas descritas aquí fueron causadas por flujos ineficientes de nutrientes terrestres y de aguas profundas a estos entornos, en general similares a los de los entornos oligotróficos en los sistemas oceánicos modernos. De hecho, una investigación geoquímica de elementos traza del Utkina Zavod y de los núcleos de perforación adyacentes de la zona de San Petersburgo sugiere que nuestras muestras se depositaron en condiciones óxicas73. Esta interpretación es apoyada independientemente por nuestros datos de las distribuciones extendidas de hopano (C31-C35) que se reducen bruscamente en abundancia por encima de los compuestos C31 con el aumento del número de carbono, lo cual es característico de la degradación de la cadena lateral de los bacteriohopanepolioles durante la diagénesis en condiciones óxicas29, y también por los bajos índices de hidrógeno, medidos por pirólisis Rock-Eval (todos inferiores a 230 mg/g de COT, y muchos están por debajo de 100 mg/g de COT) para estas muestras inmaduras debido a la formación de querógeno recalcitrante a través de la degradación oxidativa y la recondensación de la biomasa primaria pobre en lípidos en ambientes localmente óxicos (Tabla 1). Los organismos pluricelulares ediacaranos que existían en estos medios marinos poco profundos debían disponer de sustratos orgánicos suficientes para que la heterotrofia satisficiera sus necesidades de alimentación, así como de oxígeno y otros nutrientes suficientes para mantener su metabolismo. Las cuencas epicontinentales del Báltico eran probablemente más persistentemente óxicas que los entornos altamente productivos que bordeaban las zonas de oxígeno mínimo, como en la plataforma media y exterior de la cuenca salina del sur de Omán, donde la respiración de la abundante biomasa planctónica habría mantenido un oxígeno disuelto más bajo por debajo de la zona fótica.
La biota de la Ediacara y otros organismos multicelulares que viven en las costas del Báltico habrían tenido que hacer frente a fuentes de alimentación cambiantes (bacterianas frente a eucariotas) a medida que los flujos de nutrientes variaban a lo largo del tiempo, incluyendo pequeñas células y detritos orgánicos en las cuencas epicontinentales donde las bacterias eran los productores primarios dominantes (Fig. 3). Las comunidades modernas de arrecifes de coral sobreviven en entornos tropicales oligotróficos debido al eficiente reciclaje de nutrientes, incluyendo la generación de un flujo de materia orgánica disuelta (DOM) que ayuda a mantener la heterotrofia de la fauna dentro del ecosistema del arrecife74. El establecimiento de una estructura trófica marina con organismos multicelulares eucariotas sostenidos por la alimentación de detritus orgánicos tuvo que ser posterior a la expansión ambiental a escala global de los eucariotas en diversos entornos marinos, que se produjo a lo largo del intervalo Toniano-Criogénico (ca. 800-635 Ma), según se desprende de los registros de biomarcadores15. Podríamos entonces esperar que se discierna la evidencia de diferencias significativas en la comunidad marina y en la estructura trófica de una localidad a otra durante el período Ediacaran tardío, con el balance local de nutrientes seleccionando comunidades microbianas ricas en eucariotas o sin eucariotas y con la expansión progresiva de los organismos multicelulares añadiendo otra dimensión de complejidad a nivel de organismo y de comunidad. El significativo contraste regional en la disponibilidad de fosfato y otros nutrientes en los ambientes de plataforma es también una consecuencia esperada de una estructura redox marina global heterogénea para el Ediacaran tardío, antes de la oxigenación del océano profundo38.
El predominio de la productividad bacteriana del picoplancton y de los productos de degradación de la materia orgánica disuelta (DOM) asociados en las cuencas epicontinentales del Báltico podría haber sostenido un ecosistema microbiano en bucle en paralelo con la estructura trófica convencional basada en torno a productores planctónicos de mayor tamaño50. Esto puede haber favorecido diferentes modos de heterotrofia, incluyendo la alimentación en suspensión y, posiblemente, la osmotrofia, como estrategia de alimentación viable para algunos rangeomorfos, esponjas y otros organismos multicelulares del Ediacaran tardío18 junto con la emergente heterotrofia activa (móvil)19,20. Los sistemas acuáticos limitados por los nutrientes, por regla general, suelen estar dominados por el pequeño fitoplancton unicelular y el plancton heterótrofo, con bacterias que superan a los eucariotas, y que mantienen una baja biomasa neta en los entornos marinos oligotróficos del océano moderno49,51. La DOM es una fuente importante de nutrientes orgánicos y a menudo controla la productividad y la biomasa neta en los mares tropicales oligotróficos modernos, pero también lo es la co-limitación de nitrógeno y fósforo75. Del mismo modo, es probable que la DOM fuera también un sustrato importante para mantener las bacterias heterótrofas y un bucle microbiano en los antiguos entornos oligotróficos. Los tapetes microbianos bentónicos pueden haber sido un componente de esta red alimentaria dominada por bacterias, y se ha implicado a los suelos de tapetes para mejorar la preservación de la biota de Ediacara en entornos marinos7. Mientras que se encontraron valores moderados de índice de 2-metilhalfano para un subconjunto de nuestras muestras (4-10%, Tabla 1), las abundancias muy bajas de metilalcanos en relación con los n-alcanos (Fig. 1), y sólo trazas y ocurrencias esporádicas de carotenoides sugieren que los tapetes microbianos no dominaron la productividad primaria76, y que las bacterias planctónicas y, en consecuencia, sus productos de descomposición DOM florecieron. Los valores generalmente bajos del índice de hidrógeno (Tabla 1; especialmente para las muestras de Redkino), a pesar de la baja madurez térmica de los estratos y de los conjuntos de biomarcadores, son también consistentes con una entrada orgánica pobre en lípidos en entornos marginales principalmente óxicos y poco profundos. La presencia de tapetes microbianos, medida únicamente a partir de las texturas sedimentológicas, no nos dice nada sobre el equilibrio entre los eucariotas (microalgas) y las bacterias en ningún caso, ni puede limitar la contribución relativa del plancton microbiano a la productividad primaria general y al sustento de las redes alimentarias. Las comunidades de tapetes microbianos que a menudo contienen abundantes eucariotas y los carbonatos del Grupo Ara del sur de Omán con prominentes facies de laminitas trombolíticas y arrugadas son un buen ejemplo de entornos del Neoproterozoico tardío con una importante contribución de tapetes microbianos, que producen abundantes señales de esterano debido a una gran contribución de microalgas30,59.
El nivel extremadamente bajo de biomarcadores de 24-ipc sugiere que, aunque las demosponjas estaban a veces presentes, eran escasas en estos entornos. Esto podría indicar que hay menos oportunidades para los animales filtradores más pequeños que compiten con la biota Ediacara en estos entornos de baja productividad, una resistencia insuficiente de las demosponjas contra las condiciones más energéticas de las aguas poco profundas por encima de la base de las olas de buen tiempo, o una mejor adaptabilidad de las esponjas a las condiciones de bajo oxígeno mantenidas dinámicamente por debajo de la zona fótica en los entornos eutróficos (Fig. 3). En total, no se detectó 24-ipc en la mayoría de las muestras del Horizonte Redkino, que también tienen en general las relaciones H/St más elevadas de nuestro conjunto de muestras, lo que tal vez indique un cambio ecológico en la época de depósito del Horizonte Kotlin. Alternativamente, es también plausible que las esponjas habitaron estos ambientes, pero no produjeron estos biomarcadores esteroides diagnósticos en la abundancia, aunque esto parece menos probable como 24-ipc esterano está entre los compuestos de esterano C30 más comúnmente detectados en estratos de Ediacaran y aceites30,3133,59,60 y es también detectable en un subconjunto de nuestras muestras (Tabla 1).
Si bien los ambientes marinos oligotróficos persistentes sugeridos por nuestros datos representan condiciones localizadas en los océanos ediacaranos, no eran probablemente infrecuentes para las vías marinas poco profundas del Precámbrico (Fig. 3). La gran mayoría de los estudios anteriores sobre los biomarcadores ediacaranos se han llevado a cabo en rocas sedimentarias ricas en materia orgánica depositadas en entornos eutróficos y en sus productos petrolíferos, que por lo general arrojan conjuntos de biomarcadores coherentes con una contribución significativa de las fuentes microalgales44. Nuestros resultados ponen de manifiesto la importancia de estudiar una mayor variedad de entornos deposicionales, incluyendo estratos ricos en materia orgánica con una madurez térmica adecuada y diferentes litologías15,3034,59, con el fin de obtener una imagen más precisa de la escala de heterogeneidad en la química y ecología marinas de un lugar a otro. A pesar de la progresiva ventilación del océano y del aumento de la meteorización química y del suministro de nutrientes durante la ruptura de Rodinia y a lo largo del período ediacarano77,78, la heterogeneidad del océano mantuvo una variedad de condiciones químicas marinas, incluyendo ambientes pobres en nutrientes, pero habitables, que fomentaron la adaptación, la competencia y la evolución de los metazoos dentro del sistema oceánico global (Fig. 3). Mientras que las plataformas marinas eutróficas acogían a las demosponjas, pero a menudo carecían de la biota de Ediacara, siendo la cuenca salina del sur de Omán (y los gruesos afloramientos ediacaranos correlativos más al norte de Omán) un ejemplo destacado; contraintuitivamente, las vías marinas epicontinentales poco profundas, óxicas y menos productivas fueron colonizadas por la biota de Ediacara con preferencia a las demosponjas a pesar de que la DOM y otros detritos orgánicos estaban disponibles localmente para la alimentación. No se sabe si las necesidades metabólicas o la presión selectiva del medio ambiente limitaron la biota multicelular de cuerpo blando del Ediacara a estos entornos; sin embargo, nuestro estudio destaca que los entornos oligotróficos del Ediacara desempeñaron un papel potencialmente crucial en la evolución de los organismos multicelulares macroscópicos y de la ecología de las comunidades marinas.