Maturità termica eccezionalmente bassa degli strati
La maturità termica è il singolo fattore più importante che influenza la conservazione di antichi biomarcatori sedimentari, poiché le molecole organiche sono suscettibili di alterazione strutturale e stereochimica durante la progressiva sepoltura e con l’aumento dello stress termico29. Le rocce precambriane di maturità termica appropriata (maturità della finestra petrolifera o inferiore) sono prerequisiti per la conservazione di assemblaggi lipidici robusti e primari di biomarker. Molteplici rapporti di stereoisomeri di hopane e sterane, insieme a prove indipendenti da bassi valori di Tmax (per lo più all’interno di un intervallo di 417-433 °C, con una media di 426 °C) dalla pirolisi Rock-Eval, indicano che le rocce in questo studio rappresentano di gran lunga le rocce ediacarane più termicamente immature analizzate fino ad oggi utilizzando lo stato dell’arte dei metodi geochimici organici (Tabella 1). Anche se le nostre rocce hanno subito una diagenesi sedimentaria, non sono passate significativamente nella finestra petrolifera a parte i campioni di affioramento 16PL leggermente più maturi del bacino di Podillya in Moldavia, che sono ancora adatti all’analisi (maturità della finestra petrolifera medio-precoce e nessun segno evidente di contaminanti organici). La maggior parte del set allora non era stata probabilmente esposta a temperature di sepoltura superiori a 50 °C, quindi i nostri campioni sono altamente immaturi. Altre linee di evidenza molecolare, che supportano la bassa maturità termica e la sigenicità, includono (i) una dominanza di alcani policiclici biomarcatori rispetto agli n-alcani negli estratti di roccia (Fig. 2), (ii) la sopravvivenza di quantità rilevabili di 17β,21β(H)-hopanes risolvibili dai più abbondanti hopanes in possesso di configurazioni stabili 17β,21α(H)- e 17α,21β(H)-stereochemical (Tabella 1, Figure supplementari. 1 e 2), (iii) una discernibile preferenza pari-dispari tra gli n-alcani nell’intervallo da C22 a C27 (Fig. 2), poiché gli n-alcani mostrano una preferenza per il numero di carbonio solo prima della catagenesi38, e (iv) la generazione di biomarcatori di hopane e sterane termicamente immaturi dalla fase (insolubile) del cherogene mediante idrolisi catalitica (Fig. 2 supplementare). La bassa maturità termica della materia organica sedimentaria nei nostri campioni è coerente con gli indicatori di alterazione sedimentaria pubblicati in precedenza, compresi gli indici di alterazione dei conodonti e degli acritarchi, e i precedenti studi di pirolisi Rock-Eval e sui biomarcatori delle rocce sedimentarie ediacariche e paleozoiche in Baltica36,39,40, così come la mineralogia dell’argilla41. Criticamente, in ambienti in cui la maturità termica è così bassa, i meccanismi per un’alterazione significativa dei segnali primari δ15N e δ13C sono largamente assenti42.
Tabella 1 Seleziona i rapporti dei biomarcatori lipidici per la maturità termica, il biota di origine e le valutazioni ambientali deposizionali
Fig. 2
Distribuzione degli idrocarburi alifatici estraibili per un campione rappresentativo. a Cromatogramma degli ioni totali (TIC) per idrocarburi alifatici estraibili per Lugovoe #13-73 m dal Redkino Horizon. La serie n-alcani, pristane (Pr), phytane (Ph), e C27-C34 hopane (indicati dal loro numero totale di carbonio e dalla stereochimica a C-17, C-21, e C-22, ad esempio, C31αβR) sono etichettati; ** denota C29 e C30 hopenes. Si noti la dominanza dell’abbondanza di hopane su altre classi di composti alcani. b Il cromatogramma parziale degli ioni 85-Da mostra la dominanza della serie n-alcani, con una discernibile preferenza per il numero di carbonio pari e dispari nell’intervallo C22-C27, rispetto ai metilalcani
Modelli di assemblaggio di biomarcatori lipidici
Un rapporto di abbondanza dei principali (C27-C35) hopani rispetto ai principali (C27-C29) sterani è spesso usato per valutare l’equilibrio tra gli apporti di organismi di origine batterica ed eucariotica nell’antico ecosistema acquatico. Gli hopani sono fossili molecolari derivati dagli hopanoidi, che sono lipidi di membrana cellulare sintetizzati da un’ampia varietà di gruppi batterici. Allo stesso modo, gli sterani sono derivati dai precursori degli steroli, che sono prodotti quasi esclusivamente dagli eucarioti43. Hopane/sterano (H/St) rapporti per i nostri campioni coprono una gamma sorprendentemente ampia di valori da 1,6 a 119,2 (tabella 1). Per il contesto, i rapporti H/St delle rocce e degli oli neoproterozoici ricchi di elementi organici rientrano tipicamente in una gamma ristretta da 0,5 a 2,030,31,32,33. Mentre c’è una chiara differenza nei valori del più giovane Kotlin Horizon (media H/St di 8,9) e del più vecchio Redkino Horizon (media H/St di 42,9), i valori per la maggior parte dei campioni sono insolitamente alti e suggeriscono contributi anomalmente elevati di batteri.
Dal tardo Neoproterozoico, le alghe eucariotiche erano una componente ecologicamente significativa e produttori principali in molti ecosistemi marini15,34,44. La discrepanza tra il contributo globalmente importante delle alghe eucariotiche nel tardo Neoproterozoico e tuttavia i bassi livelli di biomarcatori di sterano in questi campioni indica che ci deve essere qualche determinante locale sull’abbondanza eucariotica. La spiegazione più plausibile per i rapporti hopane/sterano estremamente elevati insieme al basso contenuto di carbonio organico totale (TOC) e ai bassi indici di idrogeno (HI) trovati per i nostri campioni (Tabella 1) è che questi strati sono stati depositati in ambienti oligotrofici (cioè, fortemente limitati dai nutrienti), in cui i batteri hanno superato le alghe. Mentre è difficile trovare analoghi moderni per antichi mari epicratonici sviluppatisi durante l’alto livello del mare, osservazioni parallele di rapporti hopane/sterano più alti (fino a un ordine di grandezza) sono state trovate in precedenza per rocce sedimentarie organiche-pulite rispetto a quelle ricche di materiale organico depositate in corsi d’acqua epicontinentali ordoviciani-siluriani45.
Tipo, un basso contenuto di TOC può essere causato da una deposizione limitata di materia organica in ambienti a bassa produttività, da un basso potenziale di conservazione della materia organica nella colonna d’acqua o nei sedimenti, o dalla diluizione con un alto flusso siliciclastico. Negli oceani moderni, la produttività è più comunemente limitata da bassi livelli di nutrienti essenziali: azoto, fosforo, silicio e ferro46. L’azoto, il fosforo e il ferro sono limitati nelle regioni dell’oceano aperto dove l’upwelling, l’apporto di polvere e il deflusso costiero non forniscono sufficienti quantità di nutrienti. Nei moderni ambienti oligotrofici, il rapporto tra biomassa batterica ed eucariotica è più alto che in ambienti eutrofici o mesotrofici. È interessante notare che i campioni di affioramento Podillya 16PL dall’orizzonte Redkino della Moldavia sono fanghi contenenti fosforite e producono rapporti hopane/sterano significativamente più bassi (8.1:11.5) rispetto agli altri campioni Redkino nella tabella 1 (anche se ancora significativamente più alti di quelli trovati nel bacino salino dell’Oman meridionale o nei sedimenti fanerozoici ricchi di organico, che tipicamente rientrano in una stretta, 0.5-2.0), forse suggerendo che una maggiore disponibilità di fosfato potrebbe aver influenzato favorevolmente il rapporto eucariotico-batterico trovato localmente (dato che le firme isotopiche di azoto sono in gran parte invarianti, vedi la prossima sezione). Il contenuto di fosforo (P), così come i rapporti P/Fetotale e P/Al, sono generalmente bassi per i sedimenti del tardo Ediacarano di Podillya, Ucraina ed Estonia47 ; con l’eccezione di questo livello stratigrafico segnato da noduli di fosforite. Allo stesso modo, bassi (da ~0,01 a ~0,1 wt%) livelli di P nelle rocce siliciclastiche di Kotlin e Redkino sono stati riportati da una carota di perforazione dal margine nord-orientale della Piattaforma dell’Europa orientale11.
La topografia ampia e poco profonda nei mari epicontinentali attraverso la Baltica potrebbe aver sostenuto le limitazioni di fosforo o di altri nutrienti in ambienti marginali, a causa della precipitazione autoctona di fosfato con minerali di ferro nelle acque superficiali ossigenate e il sequestro di una serie di elementi da parte dei sedimenti shelfal48. Se il fosfato era un nutriente limitante, l’aumento del fosforo biodisponibile avrebbe aumentato la produzione primaria locale e fornito condizioni di crescita più favorevoli per il fitoplancton unicellulare di dimensioni maggiori49,50,51. Per quanto riguarda il sistema oceanico moderno, i picocianobatteri marini, Prochlorococcus e Synechococcus, sono riconosciuti per dominare il numero di cellule e la biomassa del fitoplancton negli ambienti oceanici tropicali e subtropicali oligotrofici, comprese le regioni oligotrofiche a fosfato limitato delle acque superficiali dell’oceano52. Il proclorococco e l’eterotrofo SAR11 (Pelagibacter) prosperano grazie a una serie di adattamenti, tra cui bassi costi energetici in virtù di piccoli genomi e bassi tassi di replicazione, un rapporto superficie-area-volume più elevato grazie alle dimensioni ridotte delle cellule e funzioni aggiuntive di assorbimento delle cellule per massimizzare l’utilizzo dei nutrienti53. Molti batteri sono anche in grado di sostituire i nutrienti a bassa abbondanza, ad esempio, utilizzando i solfolipidi invece dei lipidi contenenti fosforo in ambienti carenti di P54, o utilizzando substrati alternativi, ad esempio, ottenendo azoto dall’N2 atmosferico attraverso la fissazione dell’azoto55, per alleviare lo stress da nutrienti. Uno studio recente sulla sopravvivenza del batterioplancton marino in ambienti oligotrofici56 , dove il fosfato disponibile è limitato, suggerisce un ruolo importante per il metabolismo dei polifosfati negli oligotrofi marini. Gli eucarioti selezionati che competono in ambienti oligotrofici sono tipicamente piccoli picoeucarioti, che possono integrare i loro requisiti nutrizionali attraverso la mixotrofia57. Il rapporto biomassa picoeucarioti-cianobatteri tende ad aumentare in condizioni di maggiore fornitura di nutrienti49. La possibile influenza del fosforo, e altri nutrienti biolimitando, nel moderare la produttività primaria e la struttura della comunità marina in Ediacaran epeiric seaways richiede ulteriori indagini.
Senza eccezione, l’abbondanza di C29 sterani è maggiore rispetto ai corrispondenti C27 o C28 sterani per tutte le nostre località (Tabella 1). Una predominanza di C29 su C27 e C28 sterani indica probabilmente una dominanza di alghe verdi all’interno della comunità fitoplanctonica eucariotica44,58. Questa caratteristica è stata osservata nella maggior parte dei precedenti studi sui biomarcatori ediacarani15,30,31,32,33,34,35,36,44. In particolare, la distribuzione dello sterano C30 in diversi campioni di ciascuna carota di Kotlin Horizon contiene quantità basse, ma rilevabili, del biomarcatore di sterano demosponge noto come 24-isopropylcholestane (24-ipc)59,60. In totale, gli sterani 24-ipc sono stati riportati in campioni di roccia e di petrolio risalenti al Criogenio (>635 Ma fa) nel bacino salino dell’Oman meridionale e rappresentano la più antica prova di biomarcatore lipidico per i metazoi59,60. I biomarcatori 24-ipc nei nostri campioni erano circa un ordine di grandezza inferiore in abbondanza (rispetto ai rapporti totali di sterani da C27 a C30, questi erano solo 0,06-0,61%; media = 0,22%) rispetto alle rocce ediacariche e agli oli dell’Oman meridionale (1,7% in media59) o erano sotto i limiti di rilevamento a causa di abbondanza trascurabile per la maggior parte dei campioni.
Rapporti isotopici dell’azoto e del carbonio organico
Gli isotopi dell’azoto possono aiutare a discernere l’equilibrio relativo nel ciclo dell’azoto e il grado in cui la fissazione dell’azoto o la denitrificazione incompleta erano vie significative per influenzare il bilancio dei nutrienti disponibili per le comunità marine. Quando i batteri diazotrofi fissano l’azoto molecolare a causa di una mancanza di azoto fissato nella colonna d’acqua, questo può produrre valori di isotopi di azoto in massa sedimentari vicini allo 0‰61. Questo è in contrasto con le firme isotopiche positive dell’azoto (nell’intervallo da +2 a +10‰) con una modalità da +4 a +6‰ trovate per i sedimenti marini del Neoproterozoico depositati in quelle che si pensa siano condizioni di nitrato-ripieno dove il nitrato ha subito solo una parziale denitrificazione62. I valori isotopici dell’azoto per tutti i nostri campioni, ad eccezione di un valore anomalo (Tabella 1), coprono un intervallo positivo limitato da +3,5 a +6,5‰, che si sovrappone alla modalità per le rocce sedimentarie ricche di materia organica del tardo Neoproterozoico62. Vincolati da proxy redox che suggeriscono che le condizioni della colonna d’acqua ossigenata hanno prevalso, i nostri dati implicano che il nitrato ha dominato il pool di azoto inorganico disciolto. La gamma di valori δ15N suggerisce anche che N2-fissazione non era la modalità primaria di acquisizione di azoto per i produttori primari. Piuttosto, il ciclo dell’azoto era probabilmente dominato dal riciclaggio della colonna d’acqua, e l’azoto organico era soggetto a un’ossidazione quantitativa a nitrato, come avviene nelle moderne impostazioni marine prossimali. Denitrificazione bentonica e interramento N organico sarebbe stato i pozzi primari per azoto inorganico disciolto, e la gamma di valori δ15N suggerisce solo un ruolo limitato per incompleta colonna d’acqua denitrificazione, che in genere si traduce in significativo 15N-arricchimento 63. La variabilità limitata nei valori δ15N quindi probabilmente riflette δ15N di nitrato avventurato sulla piattaforma, con minore influenza dalla colonna d’acqua limitata riduzione del nitrato e N2-fissazione. Senza una forte evidenza δ15N per la fissazione dell’azoto, ipotizziamo che l’azoto non era il nutriente primario biolimite nei bacini epicontinentali del Baltica. L’isolamento da fonti fluviali ed eoliane di fosforo e le condizioni di ossidazione negli ampi bacini epicontinentali poco profondi possono aver migliorato la rimozione della fosforite autoctona e delle fasi contenenti metalli in tracce, con conseguente limitazione dei nutrienti che ha limitato la crescita e la produzione delle cellule eucariotiche.
I valori isotopici del carbonio organico totale (CTOC) vanno da -23,0 a -33,9‰, con le maggiori differenze osservate tra le diverse località dei carotaggi. L’arricchimento relativo di 13C a rapporti isotopici superiori a circa -28‰ contrasta con i dati di strati contemporanei depositati in ambienti eutrofici e marini aperti dell’Oman30,31. Tuttavia, l’intervallo che riportiamo è generalmente coerente con l’intervallo δ13CTOC per altre località della Baltica depositate nello stesso periodo di tempo11. La differenza tra Baltica e Oman potrebbe evidenziare il fatto che i valori δ13CTOC non riflettono esclusivamente un cambiamento secolare uniforme nel ciclo del carbonio durante questo intervallo di tempo31,64. Piuttosto, la gamma δ13CTOC della Baltica potrebbe, in parte, riflettere l’ecologia microbica a dominanza batterica suggerita dai rapporti unici dei biomarcatori lipidici. Le piccole dimensioni delle cellule, gli alti rapporti superficie-area-volume e i lenti tassi di crescita in condizioni oligotrofiche possono aumentare l’ampiezza del frazionamento durante l’autotrofia (εp)65,66, con conseguenti bassi valori δ13C.
Il meccanismo per le firme isotopiche più arricchite di 13C all’interno dell’intervallo riportato11 per la successione del tardo Ediacarano Baltica è meno chiaro. Il potenziale contributo della materia organica detritica, alterata dalla metamorfosi, alle successioni sedimentarie precambriane a basso TOC è un possibile meccanismo per le disparità tra i siti in δ13CTOC64. Tuttavia, il contenuto di TOC dei nostri campioni, mentre basso, è generalmente superiore a 0,10 wt%, rivelando alcuna relazione tra il contenuto di TOC e δ13C valori. Inoltre, i nostri campioni contengono materia organica termicamente immatura e modelli di biomarker lipidici che non sono coerenti con una fonte di carbonio principalmente alloctona. Più 13C-arricchito δ13CTOC firme possono essere il risultato di meccanismi alternativi per l’assimilazione del carbonio. I meccanismi di concentrazione del carbonio o l’assorbimento attivo del bicarbonato da parte dei procarioti possono risultare in valori più piccoli per εp e valori δ13CTOC più alti67,68. L’emergere del biota ediacariano può aver ampliato significativamente le reti alimentari marine e stimolato nuove vie di eterotrofia microbica, compresi possibili contributi dal complesso ciclo del carbonio all’interno dei tappeti microbici bentonici. Con un’ulteriore considerazione per il ruolo potenzialmente importante della materia organica disciolta69 come fonte di carbonio, l’ampiezza dei valori δ13CTOC riflette probabilmente una gamma di processi biogeochimici di ciclo del carbonio che potrebbero essere unici per l’ambiente marino ediacariano in evoluzione e potrebbero essere legati a un fenomeno per il quale non abbiamo analoghi moderni rappresentativi.
Gli antichi assemblaggi di biomarcatori lipidici e gli indicatori di isotopi stabili per la produzione bentonica di stuoie microbiche in tandem con la limitazione dell’azoto fisso sono stati trovati nelle rocce del primo Triassico della Cina meridionale70. All’indomani dell’estinzione di massa di fine Permiano a Meishan, c’è stato un grande picco nei rapporti hopane/sterano (fino a circa 60) accompagnato da un forte spostamento nella firma isotopica Norg a valori da 0 a -2‰, coerente con la diazotrofia batterica. Questo è associato a un alto segnale di 2-metil-opani (indice 2-metil-opani fino al 33%) e metilalcani distintivi che indicano la proliferazione di tappeti microbici bentonici. Non osserviamo una tendenza simile nella firma isotopica dell’azoto o nei modelli di biomarcatori che ovviamente indicherebbero un significativo apporto microbico all’interno del nostro set di dati ediacarani, anche se un singolare valore di 15N impoverito è stato trovato nei nostri campioni (Tabella 1). Pertanto, il forte segnale batterico osservato nei nostri campioni Baltica probabilmente non è prevalentemente una firma di stuoie microbiche bentoniche.
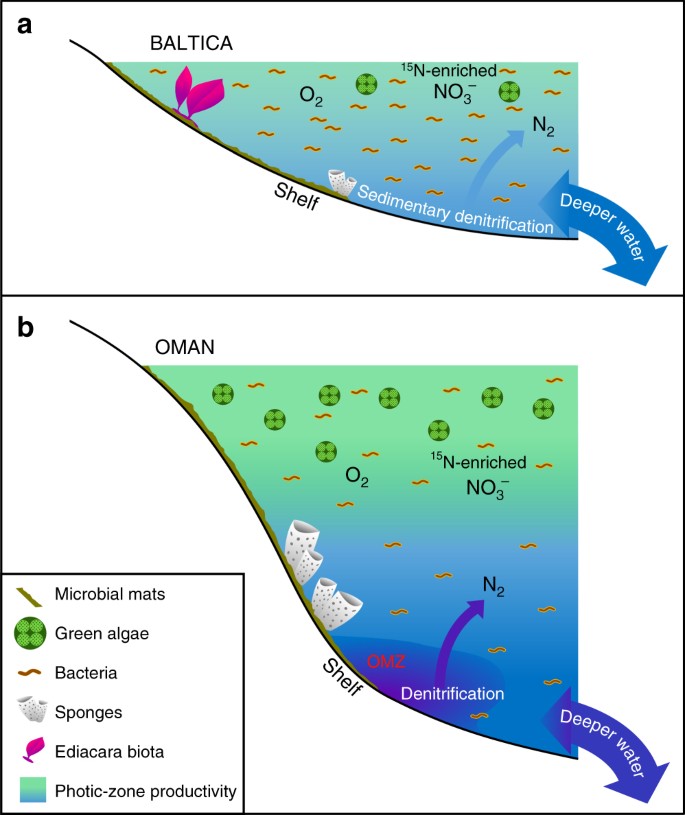
Sostentamento paleoambientale del biota di Ediacara vs. Le condizioni apparentemente oligotrofiche attraverso i bacini epicratonici e i margini continentali della Baltica, mentre si spostava dalle alte alle basse latitudini dal tardo Ediacarano all’inizio del Cambriano, potrebbero essere associate sia all’avvezione limitata di acque profonde relativamente ricche di nutrienti, sia all’esaurimento dei nutrienti, derivante dall’assimilazione e dalla pulizia durante il trasporto e la deposizione attraverso questi ampi bacini epicontinentali poco profondi (Fig. 3). Questi bacini sono stati episodicamente isolati dagli oceani e hanno sviluppato condizioni di ipersalinità (ad esempio, durante il tempo di Redkino) e salmastre (ad esempio, durante il tempo di Kotlin71). La stabilità tettonica a lungo termine ha portato a una topografia a basso rilievo della Baltica del tardo Ediacarano, altamente suscettibile alle inondazioni e a una fornitura inefficiente di fosforo derivato dagli agenti atmosferici. In contrasto con la Baltica, gli studi sui biomarcatori degli strati del supergruppo Huqf in Oman hanno rivelato un ecosistema eutrofico, ricco di microalghe30,31,59, ma privo di biota Ediacara, anche in estesi affioramenti nelle montagne dell’Oman e nella regione dell’Huqf, nelle impostazioni da interno a esterno della piattaforma3. La paleogeografia del bacino salino dell’Oman meridionale è stata ricostruita per il tardo Neoproterozoico a ~13° dall’equatore nell’emisfero meridionale, ampiamente simile alla paleolatitudine di Baltica72.
Fig. 3
Maggiori differenze tra ambienti marini ediacarani a bassa produttività e produttivi. I diagrammi schematici sono mostrati per a estesi margini di bacini epicontinentali oligotrofici e poco profondi della Baltica spesso dominati dalla produttività batterica dove la fauna molle di Ediacara fiorì e dove la denitrificazione e l’anammox probabilmente erano limitati ai sedimenti; e b impostazioni di piattaforma eutrofica e più profonda del bacino salino dell’Oman meridionale, dove le alghe verdi prosperavano come produttore primario e le demospugne erano abbondanti, ma la fauna molle di Ediacara non era prominente (con fossili di biota di Ediacara assenti anche in affioramenti ediacarani correlati nell’Oman settentrionale). Sui margini continentali produttivi, la denitrificazione e l’anammox si sono probabilmente verificati sia nella colonna d’acqua che nei sedimenti
Non ci sono prove di anossia persistente ed estesa sui margini continentali poco profondi della Baltica durante il tardo Ediacarano11, e suggeriamo che le condizioni oligotrofiche qui descritte sono state causate da inefficienti flussi di nutrienti terrestri e di acque profonde a queste impostazioni, ampiamente simili a quelli in ambienti oligotrofici nei sistemi oceanici moderni. Infatti, un’indagine geochimica degli elementi in traccia del Utkina Zavod e delle carote di perforazione adiacenti dell’area di San Pietroburgo suggerisce che i nostri campioni sono stati depositati in condizioni di oxic73. Questa interpretazione è supportata indipendentemente dai nostri dati dalle distribuzioni estese di hopane (C31-C35) che scendono bruscamente in abbondanza sopra i composti C31 con l’aumento del numero di carbonio, che è caratteristico della degradazione della catena laterale dei batteriohopanepolyols durante la diagenesi in condizioni ossessive29, e anche da indici di basso idrogeno, come misurato dalla pirolisi Rock-Eval (tutti inferiori a 230 mg/g TOC, e molti sono inferiori a 100 mg/g TOC) per questi campioni immaturi dovuti alla formazione di cherogene recalcitrante attraverso la degradazione ossidativa e la ricondensazione della biomassa primaria povera di lipidi in ambienti localmente tossici (Tabella 1). Gli organismi multicellulari dell’Ediacarano che esistevano in questi ambienti marini poco profondi dovevano avere sufficienti substrati organici per l’eterotrofia per soddisfare le loro esigenze di alimentazione insieme a sufficiente ossigeno e altri nutrienti per sostenere il loro metabolismo. I bacini epicontinentali della Baltica erano probabilmente più persistentemente ossigenati degli ambienti altamente produttivi che circondavano le zone minime di ossigeno, come sulla piattaforma media ed esterna del bacino salino dell’Oman meridionale, dove la respirazione dell’abbondante biomassa planctonica avrebbe mantenuto l’ossigeno disciolto più basso sotto la zona fotica.
Il biota Ediacara e altri organismi multicellulari che vivono al largo della Baltica avrebbero dovuto far fronte a fonti di cibo mutevoli (batteriche vs. eucariotiche) come i flussi di nutrienti variavano nel tempo, tra cui piccole cellule e detriti organici in bacini epicontinentali dove i batteri erano i produttori primari dominanti (Fig. 3). Le moderne comunità della barriera corallina sopravvivono in ambienti tropicali oligotrofici grazie all’efficiente riciclo dei nutrienti, compresa la generazione di un flusso di materia organica disciolta (DOM) che aiuta a sostenere l’eterotrofia faunistica all’interno dell’ecosistema della barriera74. L’istituzione di una struttura trofica marina con organismi multicellulari eucarioti sostenuti dall’alimentazione su detriti organici doveva postdatare l’espansione ambientale su scala globale degli eucarioti in diversi ambienti marini, che si è verificata attraverso l’intervallo Toniano-Criogenico (circa 800-635 Ma), come misurato da record di biomarker15. Potremmo quindi aspettarci di discernere l’evidenza di differenze significative nella comunità marina e nella struttura trofica da località a località durante il tardo Ediacarano, con l’equilibrio dei nutrienti locale che seleziona le comunità microbiche ricche o prive di eucarioti e con la progressiva espansione degli organismi multicellulari che aggiunge un’altra dimensione di complessità a livello di organismi e comunità. Un contrasto regionale significativo nella disponibilità di fosfato e di altri nutrienti negli ambienti di piattaforma è anche una conseguenza attesa di una struttura redox marina globale eterogenea per il tardo Ediacaran, prima dell’ossigenazione dell’oceano profondo38.
La dominanza della produttività batterica del picoplancton e dei prodotti di degradazione della materia organica disciolta (DOM) associati nei bacini epicontinentali della Baltica potrebbe aver sostenuto un ecosistema microbico in parallelo con la struttura trofica convenzionale basata su produttori planctonici di grandi dimensioni50. Questo potrebbe aver favorito diverse modalità di eterotrofia, tra cui l’alimentazione in sospensione e, possibilmente, l’osmotrofia, come strategia di alimentazione praticabile per alcuni rangeomorfi, spugne e altri organismi multicellulari del tardo Ediacarano18 insieme all’eterotrofia attiva (motile) emergente19,20. I sistemi acquatici limitati dai nutrienti, come regola generale, sono spesso dominati da piccolo fitoplancton unicellulare e da plancton eterotrofo, con batteri che superano gli eucarioti e sostengono una bassa biomassa netta in ambienti marini oligotrofi dell’oceano moderno49,51. Il DOM è un’importante fonte di nutrienti organici e spesso controlla la produttività e la biomassa netta nei moderni mari tropicali oligotrofici, ma anche la co-limitazione di azoto e fosforo75. Allo stesso modo, il DOM era probabilmente un substrato importante per sostenere i batteri eterotrofi e un ciclo microbico nelle antiche impostazioni oligotrofe. Le stuoie microbiche bentoniche possono essere state una componente di questa rete alimentare dominata dai batteri, e i terreni delle stuoie sono stati implicati per migliorare la conservazione del biota di Ediacara in ambienti marini7. Mentre valori moderati di 2-methylhopane indice sono stati trovati per un sottoinsieme dei nostri campioni (4-10%, Tabella 1), abbondanze molto basse di metilalcani rispetto a n-alcani (Fig. 1), e solo tracce e sporadiche occorrenze di carotenoidi suggeriscono che tappeti microbici non ha dominato la produttività primaria 76, e che i batteri planctonici e, corrispondentemente, i loro prodotti di degradazione DOM fiorito. I valori generalmente bassi dell’indice di idrogeno (Tabella 1; specialmente per i campioni di Redkino), nonostante la bassa maturità termica degli strati e degli assemblaggi di biomarker, sono anche coerenti con l’apporto organico povero di lipidi in ambienti marginali principalmente tossici e poco profondi. Le presenze di stuoie microbiche misurate solo dalle strutture sedimentologiche non ci dicono nulla sull’equilibrio tra eucarioti (microalghe) e batteri in ogni caso, né possono vincolare il contributo relativo del plancton microbico alla produttività primaria complessiva e al sostentamento delle reti alimentari. Le comunità microbiche della stuoia che spesso contengono eucarioti abbondanti e i carbonati del gruppo Ara dall’Oman del sud con facies trombolitiche e laminitiche prominenti sono un buon esempio di ambienti del tardo Neoproterozoico con un significativo contributo microbico della stuoia, producendo abbondanti segnali sterani dovuti a un grande contributo di microalghe30,59.
Il livello estremamente basso di biomarcatori 24-ipc suggerisce che mentre i demosponges erano talvolta presenti, essi erano radi in questi ambienti. Questo potrebbe indicare minori opportunità per i piccoli animali che si nutrono di filtri in competizione con il biota Ediacara in questi ambienti a bassa produttività, insufficiente resilienza delle demospugne contro le condizioni più energetiche di mare poco profondo sopra la base delle onde fair-weather, o migliore adattabilità delle spugne alle condizioni di basso ossigeno mantenute dinamicamente sotto la zona fotica in ambienti eutrofici (Fig. 3). In totale, 24-ipc non è stato rilevato nella maggior parte dei campioni del Redkino Horizon, che hanno anche generalmente i più alti rapporti H/St tra il nostro set di campioni, forse indicando un cambiamento ecologico dal tempo deposizionale Kotlin Horizon. In alternativa, è anche plausibile che le spugne abbiano abitato questi ambienti, ma non abbiano prodotto questi biomarcatori steroidei diagnostici in abbondanza, anche se questo sembra meno probabile in quanto lo sterano 24-ipc è tra i composti steranici C30 più comunemente rilevati negli strati ediacarani e negli oli30,3133,59,60 ed è anche rilevabile in un sottoinsieme dei nostri campioni (Tabella 1).
Mentre gli ambienti marini oligotrofici persistenti suggeriti dai nostri dati rappresentano condizioni localizzate negli oceani ediacarani, probabilmente non erano rari per i corsi d’acqua precambriani poco profondi (Fig. 3). La stragrande maggioranza dei precedenti studi biomarcatore Ediacaran sono stati condotti su rocce sedimentarie ricche di organico depositate in ambienti eutrofici e loro prodotti petroliferi, che in genere producono assemblaggi biomarcatore coerente con il contributo significativo fonte microalgale 44. I nostri risultati evidenziano l’importanza di studiare una più ampia varietà di ambienti deposizionali, tra cui strati ricchi di materia organica di maturità termica appropriata e litologie diverse15,3034,59, al fine di ottenere un quadro più preciso per la scala di eterogeneità nella chimica ed ecologia marina da luogo a luogo. Nonostante la progressiva ventilazione dell’oceano e l’aumento dell’erosione chimica e dell’apporto di nutrienti durante la disgregazione della Rodinia e per tutto il periodo ediacariano77,78, l’eterogeneità dell’oceano ha mantenuto una varietà di condizioni chimiche marine, compresi ambienti poveri di nutrienti ma abitabili che hanno favorito l’adattamento, la competizione e l’evoluzione dei metazoi nel sistema oceanico globale (Fig. 3). Mentre le piattaforme marine eutrofiche ospitavano demospugne, ma spesso mancavano del biota Ediacara, con il bacino salino dell’Oman meridionale (e i correlati spessi affioramenti ediacarani più a nord in Oman) che ne sono un esempio importante; controintuitivamente, i corsi d’acqua epicontinentali poco profondi, ossigenati e meno produttivi erano colonizzati dal biota Ediacara di preferenza alle demospugne nonostante il DOM e altri detriti organici fossero disponibili localmente per l’alimentazione. Se i requisiti metabolici o la pressione selettiva ambientale hanno ristretto il biota multicellulare molle Ediacaran a queste impostazioni rimane incerto; tuttavia, il nostro studio evidenzia che le impostazioni oligotrofiche Ediacaran hanno giocato un ruolo potenzialmente cruciale nell’evoluzione degli organismi multicellulari macroscopici e nell’ecologia della comunità marina.