Exceptionally low thermal maturity 地層の
熱的成熟度は、古代の堆積バイオマーカーの保存に影響を与える最も重要な要因である。なぜなら、有機分子は埋没が進行し、熱応力が増加すると、構造および立体化学的に変化しやすくなるからである29。 適切な熱成熟度(オイルウィンドウ成熟度以下)の先カンブリア岩石は、強固で主要なバイオマーカーの脂質群を保存するための前提条件である。 複数のホパンとステランのバイオマーカーの立体異性体比、および Rock-Eval 熱分解による低い Tmax 値(ほとんどが 417-433 ℃の範囲、平均 426 ℃)から独立した証拠により、本研究の岩石は、最先端の有機地球化学的手法でこれまで分析したエディアカラ岩石の中で、最も熱的に未成熟であることがわかった(表 1)。 我々の岩石は堆積斜行を受けたが,モルドバのポディリア盆地から採取したやや成熟した16PL露頭試料を除けば,オイルウィンドウに大きく移行しておらず,まだ分析に適している(オイルウィンドウ初期〜中期成熟で有機汚染物質の明らかな兆候はない). また,このサンプル群の大部分は,50℃を超える埋没温度にさらされたことがないため,非常に未熟なサンプルであると考えられる。 低熱成熟度と合成性を裏付ける他の分子的証拠としては、(i) 岩石抽出物中のn-アルカンよりも多環系バイオマーカーアルカンが優勢であること (Fig. 2)、(ii) より豊富で安定な17β,21β(H)-および17α,21β(H)-立体化学構成を持つホパンと分離可能な検出量のホパンの生存 (Table 1、補足図.) が挙げられ、これらは、低熱成熟度と合成性の証拠である。 また、(iv) 触媒的加水分解により、不溶性ケロジェン相から熱的に未成熟なホパンおよびステランのバイオマーカーが生成された(補図2)。 我々の試料における堆積有機物の低い熱成熟度は、コノドントやアクリタークの変質指標、バルト海のエディアカラや古生代の堆積岩の過去のRock-Eval熱分解・バイオマーカー研究36,39,40、および粘土鉱物学41などの既往の堆積物変質の指標と整合的であった。 決定的なのは、熱成熟度がこれほど低い環境では、一次δ15Nとδ13Cシグナルの著しい変質のメカニズムはほとんど存在しない42.

代表サンプルの抽出可能脂肪族炭化水素の分布。 a Redkino HorizonのLugovoe #13-73 mの抽出可能脂肪族炭化水素のトータルイオンクロマトグラム (TIC). n-アルカン系列、プリスタン(Pr)、フィタン(Ph)、C27-C34ホパン(総炭素数とC-17、C-21、C-22の立体化学で表記、例:C31αβR)、**はC29およびC30ホペンのラベル付き。 他のアルカン化合物よりもホパン化合物の方が多いことに注意。 b 部分的な85-Daイオンクロマトグラムでは、n-アルカンシリーズが優勢であり、C22-C27の範囲では、奇数-偶数の炭素数の優先性が明瞭に見られる。 9143>
Lipid biomarker assemblage patterns
Abundance ratio of major (C27-C35) hopanes to major (C27-C29) steranes are often used to assess balance of bacterial versus eukaryotic source organism inputs to ancient aquatic ecosystem.A B.
Absisting of Biological in a rapid rapid rapid rapid rapid level. ホパン類は、多種多様な細菌群が合成する細胞膜脂質であるホパノイドに由来する分子化石である。 同様に、ステランはステロール前駆体に由来し、真核生物によってほぼ独占的に生産される43。 我々の試料のホパン/ステラン(H/St)比は、1.6から119.2まで、驚くほど大きな範囲を占めている(表1)。 有機物に富む新第三紀の岩石やオイルのH/St比は、通常0.5から2.030,31,32,33の狭い範囲に収まっている。 若いKotlin Horizon(平均H/St 8.9)と古いRedkino Horizon(平均H/St 42.9)の値には明確な違いがあるが、ほとんどのサンプルの値は異常に高く、バクテリアの寄与が異常に高いことを示唆している。
新生代後半までに、真核藻類は多くの海洋生態系の重要な構成要素と主要生産者となっていた15,34,44。 新第三紀後期における真核藻類の世界的に重要な貢献と、これらの試料における低レベルのステランバイオマーカーとの間の不一致は、真核生物の存在量に何らかの局所的な決定要因があるはずであることを示すものである。 我々の試料(表1)で見つかった、低い全有機炭素量(TOC)と低い水素指数(HI)と共に極めて高いホパン/ステラン比の最も簡潔な説明は、これらの地層が貧栄養環境(すなわち、強い栄養制限)で堆積し、その中でバクテリアが藻類と競争していたというものである。 高海水準の時代に形成された古代のエピクラトン海を現代に再現することは困難であるが、オルドビス紀からシルル紀の大陸外海に堆積した有機物に富んだ堆積岩に対して、ホパン/ステラン比が高い(最大で1桁の差がある)ことが、以前に平行して観察されている45。
一般的に、低いTOC含有量は、低生産性環境における有機物の限られた堆積、水柱または堆積物における有機物の低い保存性、または高い珪質フラックスによる希釈によって引き起こされることがある。 現代の海洋では、生産性は、窒素、リン、ケイ素、鉄といった必須栄養素の低レベルによって最もよく制限されている46。 窒素、リン、鉄は、湧昇、ダスト投入、沿岸流出が十分な量の栄養塩を供給しない外洋域で制限される。 現代の貧栄養環境では、真核生物に対するバクテリアのバイオマスの比率が、富栄養環境や中栄養環境よりも高くなる。 興味深いことに、モルドバの Redkino 地層の Podillya 16PL 露頭試料はリン酸塩を含む泥岩で、表 1 の他の Redkino 試料よりもかなり低いホパン/ステラン比(8.1:11.5)が得られた(それでも南オマーン塩盆や有機物に富む花生層堆積物で見られるものよりかなり高く、通常 0.5 という狭い範囲に収まっている)。5-2.0の範囲にある)、おそらくリン酸塩の利用可能性が増加し、その地域で見られる真核生物と細菌の比率に好影響を与えた可能性を示唆している(窒素同位体標識がほぼ一定であることを考えると、次のセクションを参照)。 ポディリア、ウクライナ、エストニアの後期エディアカラ堆積物では、リン(P)含有量、P/Fetotal および P/Al 比は概して低く、リン鉱石ノジュールのあるこの層準を例外とする47。 同様に、East European Platform の北東縁のドリルコアから、Kotlin と Redkino の珪長質岩中の P が低い(~0.01~~0.1 wt%)ことが報告されている11。
バルト海の広い浅い地形は、酸化的な表面水中の鉄鉱物とともにリン酸塩の自己沈殿と棚状堆積物による様々な元素の隔離により、限界環境におけるリンまたは他の栄養塩の制限を維持した可能性がある48。 もしリン酸塩が制限的な栄養素であったならば、生物学的利用可能なリンが増加すれば、地域の一次生産が促進され、より大きなサイズの単細胞植物プランクトンにとってより好ましい成長条件が提供されたであろう49,50,51。 現代の海洋システムでは、海洋性ピコシアノバクテリアであるプロクロロコッカスとシネコッカスが、海洋表層水のリン酸が制限された貧栄養領域を含む貧栄養熱帯・亜熱帯海洋環境において、植物プランクトンの細胞数およびバイオマスを支配していると認識されている52。 プロクロロコッカスや従属栄養生物であるSAR11(ペラジバクター)は、ゲノムが小さく複製率が低いことによる低エネルギーコスト、細胞サイズが小さいことによる高い表面積対体積比、栄養素を最大限に利用するための細胞取り込み機能の付加などの様々な適応性によって繁栄することができる53。 また、多くの細菌は、例えば、リン欠乏環境においてリンを含む脂質の代わりにスルホリピドを利用することで低存在量の栄養素を代替したり54、窒素固定によって大気中の窒素を調達するなど55、代替基質を利用することで栄養ストレスを緩和することが可能である。 利用可能なリン酸が限られた貧栄養環境における海洋細菌プランクトンの生存に関する最近の研究56は、海洋貧栄養生物におけるポリリン酸代謝の重要な役割を示唆している。 貧栄養環境で競争する真核生物は、通常、小型のピコユーカリオテであり、ミクソトロフィーによって栄養所要量を補っている可能性がある57。 ピコユーカリオテとシアノバクテリアのバイオマス比は、栄養供給が増加した場合に増加する傾向がある49。 エディアカラの大平洋海域における基礎生産性や海洋生物群集の構造を調整する上で、リンやその他の生物制限栄養素の影響を受ける可能性については、さらなる調査が必要である。 C29ステランがC27やC28ステランより多いということは、真核植物プランクトン群集の中で緑藻類が優勢であることを示していると思われる44,58。 この特徴は、これまでのエディアカランのバイオマーカー研究のほとんどで観察されている15,30,31,32,33,34,35,36,44. 注目すべきは、Kotlin Horizon の各ドリルコアから採取したいくつかのサンプルの C30 ステラン分布に、24-イソプロピルコレスタン (24-ipc) として知られるデモスポンジステランバイオマーカーが低いが検出できる量含まれていることである59, 60. 24-ipc ステランは、南オマーン塩盆の低温期(>635 Ma 前)の岩石と油のサンプルで報告されており、後生動物に関する最も古い脂質バイオマーカーである59,60。 我々の試料中の24-ipcバイオマーカーは、南オマーンのエディアカラ岩石やオイル(平均1.7%59)と比較して、存在量が1桁程度低いか(C27からC30ステラン比に対して、これらはわずか0.06から0.61%;平均0.22%)、大部分の試料で存在量がごくわずかで検出限界以下であった。
窒素と有機炭素の同位体比
窒素同位体は、窒素循環の相対的なバランス、および窒素固定または不完全な脱窒が海洋社会で利用できる栄養バランスに影響を与える重要な経路だった程度を見分けるのに役立ちます。 水柱に固定窒素がないため、ジアゾテクノロジー細菌が分子状窒素を固定すると、堆積物のバルク窒素同位体比は0‰61に近い値を示すことがある。 これは、硝酸塩が部分的にしか脱窒されていないと考えられる硝酸塩充足条件下で堆積した新原生代の海洋堆積物に見られる正の窒素同位体比(+2〜+10‰の範囲)、+4〜+6‰のモードとは対照的である62。 1 つの異常値を除くすべての試料(表 1)の窒素同位体比は +3.5 から +6.5 ‰の限られた正の範囲にあり、これは新生代後期の有機物に富む堆積岩のモードと重なる62。 水柱が酸化的であったことを示唆する酸化還元プロキシによって制約され、我々のデータは硝酸塩が溶存無機窒素プールを支配していたことを示唆している。 また、δ15Nの値の範囲は、N2固定が一次生産者の窒素獲得の主要な様式でなかったことを示唆している。 むしろ、窒素循環は水柱のリサイクルによって支配されていたと思われ、有機窒素は現代の近海環境と同様に硝酸塩への定量的酸化を受けた。 溶存無機窒素の吸収源は底生動物の脱窒と有機窒素の埋没であり、δ15N 値の範囲は、通常著しい 15N 濃縮をもたらす不完全な水柱の脱窒の役割が限られていることを示唆している63 。 したがって、δ15N値の限られた変動は、プラットフォームに移流された硝酸塩のδ15Nを反映していると考えられ、水柱の限られた硝酸塩還元とN2固定による影響はわずかであると思われる。 窒素固定に関する強いδ15Nの証拠がないため、バルト海の大陸棚流域では窒素は主要な生物制限栄養素ではなかったと考えられる。 9143>
全有機炭素(CTOC)同位体比は-23.0から-33.9‰の範囲で、異なるドリルコア地点間で最も大きな差が観察された。 同位体比が約-28‰より高い場合の相対的な13C濃縮度は、オマーンの富栄養開放環境において堆積した同時期の地層から得られたデータと対照的である30,31。 しかし、我々が報告した範囲は、同時期に堆積したバルティカの他の場所のδ13CTOC範囲と概ね一致する11。 バルチカとオマーンの違いは、δ13CTOC 値がこの時期の炭素循環の一様な経年変化を排他的に反映していないことを強調しているのかもしれない31,64。 むしろ、バルチカからのδ13CTOCの範囲は、ユニークな脂質バイオマーカー比が示唆するバクテリア優位の微生物生態を一部反映しているのかもしれない。 小さな細胞サイズ、高い表面積対体積比、および貧栄養条件下での遅い成長速度は、自家増殖中の分画の大きさ(εp)65,66を増加させ、低いδ13C値をもたらす。
後期エディアカラ紀バルチカ連続体の報告11範囲内でより13C濃縮同位体の署名のメカニズムは、あまり明確になっていない。 先カンブリア時代の低TOC堆積物サクセッションにおいて,デトリタルの変成有機物が寄与している可能性は,δ13CTOC64のサイト間格差のメカニズムとして考えられる. しかし、我々の試料の TOC 含有量は低いものの、概ね 0.10 wt% 以上であり、TOC 含有量と δ13C 値の関係は明らかにされていない。 さらに、我々の試料は熱的に未熟な有機物を含んでおり、バイオマーカーの脂質パターンは主にアロキソニアス炭素源とは矛盾するものであった。 より13Cに富むδ13CTOCシグネチャーは、炭素同化のための別のメカニズムの結果である可能性がある。 原核生物による炭素濃縮機構や重炭酸塩の積極的な取り込みは、εpの値を小さくし、δ13CTOCの値を大きくすることができる67,68。 エディアカラ生物群の出現は海洋食物網を著しく拡大し、底生微生物マット内の複雑な炭素循環による貢献を含む、微生物の異栄養の新しい道を刺激した可能性がある。 さらに、溶存有機物69の炭素源としての重要な役割を考慮すると、δ13CTOC値の幅は、進化するエディアカラ海洋環境に特有の生物地球化学的炭素循環プロセスの範囲を反映していると考えられ、現代の代表的な類似物質がない現象に関連している可能性がある。
中国南部の初期三畳紀の岩石から、固定窒素の制限と同時に底生生物、微生物マット生産のための古代の脂質バイオマーカー集合と安定同位体指標が発見されている70。 梅山では、ペルム紀末の大量絶滅の余波で、ホパン/ステラン比の大きなスパイク(最大約60)が見られ、それに伴ってNorg同位体比が0〜-2‰の値に強くシフトし、細菌のジアゾ栄養と一致した。 これは、2-メチルホパン(2-メチルホパン指数33%まで)の高い信号と底生微生物マットの増殖を示す特徴的なメチルアルカンと関連している。 我々のエディアカランのデータセット内では、特異な15N欠乏値が見つかったものの、明らかに著しい微生物マットの投入を指し示すような窒素同位体比シグネチャーやバイオマーカーパターンに同様の傾向は見られない(表1)。 そのため,Baltica試料で観測された強い細菌シグナルは,底生微生物マットのシグネチャーとして優勢ではない可能性が高い.
Ediacara biota vs. demosponges
Ediacaran 後期からカンブリア初期に高緯度から低緯度へ移動したバルティカのエピクラトンおよび大陸縁盆地全域に見られた貧栄養状態は、比較的栄養豊富な深海水の移流制限または、これらの広く浅海のエピ大陸盆地にわたる輸送および堆積中の同化および清掃による栄養枯渇と関連があるかもしれない(図3)。 これらの盆地は一時的に海洋から隔離され、過塩分(例:レッドキノ時代)および汽水域(例:コトリン時代71)が形成された。 長期にわたるテクトニックの安定性により、後期エディアカラ期のバルティカは低レリーフの地形となり、洪水の影響を強く受け、風化由来のリンの供給が非効率的になった。 Balticaとは対照的に,オマーンのHuqf超群の地層を用いたバイオマーカー研究では,オマーン山地やHuqf地域の内棚から外棚にかけての広範囲な露頭でも,微細藻類に富む富栄養化した生態系30, 31, 59が存在したが,エディアカラ生物相を欠いていた3. 南オマーン塩盆の古地理は,南半球の赤道から〜13°で新第三紀後期に復元され,バルチカ72古緯度にほぼ近い。
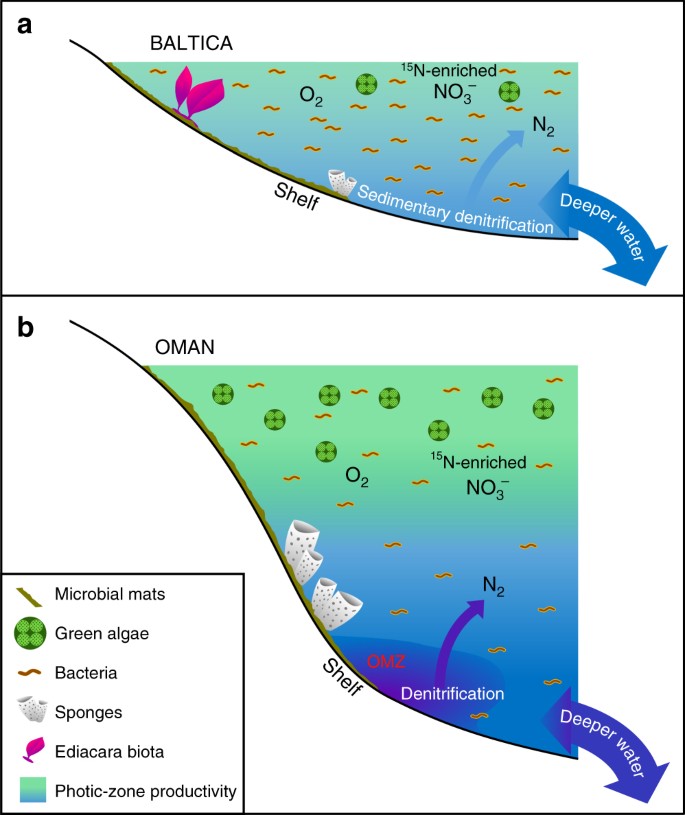
低生産性と生産性のエディアカラ海洋環境における大きな相違点. バルト海の広範な貧栄養で浅い海盆の縁では,バクテリア生産性が支配的で,エディアカラ軟体動物が繁栄し,脱窒とアナモックスが堆積物に限定されていたと思われる模式図が示される。 b 南オマーン塩水盆の富栄養で深海の棚では、一次生産者として緑藻類が繁栄し、海綿動物も豊富だったが、エディアカラ軟体動物相は顕著ではなかった(オマーン北部の相関するエディアカラ露頭ではエディアカラ生物相化石も欠落している)。 生産性の高い大陸縁では、水柱と堆積物の両方で脱窒とアナモックスが起こったと考えられる
エディアカラ後期にバルティカの浅い大陸縁とその周辺で持続的かつ広範囲な無酸素状態を証明するものはない11.Baltica(バルティカ)大陸縁の堆積物(珪藻土)層が、エディアカラ紀後期に珪藻土を含有しないことがわかった。 そして、ここで述べた貧栄養状態は、現代の海洋システムにおける貧栄養環境とほぼ同様に、これらの環境における陸上および深海の栄養フラックスの非効率性によって引き起こされたことを示唆する。 実際、サンクトペテルブルグ地域のUtkina Zavodと隣接するドリルコアの微量元素の地球化学的調査は、我々のサンプルが酸化的条件下で堆積したことを示唆している73。 この解釈は、拡張ホパン (C31-C35) 分布が、炭素数の増加とともに C31 化合物よりも存在量が急激に減少することから、酸化条件下でのダイアジェネシスにおけるバクテリオホパンポリオールの側鎖分解に特徴的なデータからも独立して裏付けられている29。 また、Rock-Eval 熱分解で測定された低水素指数(すべて 230 mg/g TOC 以下、多くは 100 mg/g TOC 以下)により、これらの未成熟な試料は、局所的な酸化環境において脂質の少ない一次バイオマスの酸化分解と再凝縮により難溶性のケロジェンが形成されたためと考えられる (Table 1)。 このような浅海域に生息するエディアカランの多細胞生物は、餌となる有機基質と代謝を維持するための酸素や栄養塩を十分に有していたものと思われる。 バルチカの大陸棚盆地は、酸素極小帯に縁取られた生産性の高い環境、例えば南オマーン塩盆の中層から外層にかけて、より持続的に酸化的であった可能性が高い。
バルト海沖に生息するエディアカラ生物群やその他の多細胞生物は、バクテリアが主要な一次生産者であった大陸棚盆地の小細胞や有機デトリタスなど、栄養フラックスが時代によって変化する中で、餌源(バクテリア対真核生物)の変化に対処する必要があったであろう(図3)。 現代のサンゴ礁生態系は、熱帯の貧栄養環境においても、溶存有機物(DOM)フラックスの生成を含む効率的な栄養塩のリサイクルにより、サンゴ礁生態系内の動物種の異栄養を維持することで生存している74。 真核生物の多細胞生物が有機デトリタスを餌として維持される海洋栄養構造が確立されたのは、トニアン-クリオジェニアン期(約800〜635Ma)に起こった真核生物の多様な海洋環境への地球規模の環境拡大の後でなければならなかったとバイオマーカーの記録から判断される15。 エディアカラ紀後期には、地域によって海洋生物群集や栄養構造に大きな違いがあり、地域の栄養バランスが真核生物に富む微生物群集や真核生物に乏しい微生物群集を選択し、多細胞生物の漸増が生物レベルや群集レベルで別の次元の複雑さを増している証拠が発見されると期待できるかもしれない。
バルティカ大陸上盆地におけるピコプランクトン細菌生産力とそれに伴う溶存有機物(DOM)分解産物の支配は、大型プランクトン生産者周辺に基づく従来の栄養構造と並行して、微生物ループエコシステムを維持した可能性がある50。 このことは、浮遊性摂食や、おそらくは浸透性摂食を含む異なる様式の異栄養が、一部のレンジモルフや海綿動物、その他のエディアカラ後期の多細胞生物18にとって、新たな活動性(運動性)異栄養と合わせて、実行可能な摂食戦略として好まれたのかもしれない19, 20. 一般的に栄養不足の水系は、小さな単細胞植物プランクトンや従属栄養プランクトンが多く、バクテリアは真核生物に勝り、現代海洋の貧栄養海洋環境では低い純バイオマスを維持する49,51。 DOMは重要な有機栄養源であり、現代の貧栄養熱帯海域の生産性と純バイオマスをしばしば制御するが、窒素とリンの共限界も同様である75。 同様に、DOMもまた、古代の貧栄養環境において従属栄養細菌と微生物ループを維持するための重要な基質であったと思われる。 海底の微生物マットは、この細菌が支配する食物網の構成要素であった可能性があり、マット地は海洋環境におけるエディアカラ生物群の保存を促進することが示唆されている7。 2-メチルホパンインデックスが一部のサンプルで中程度の値(4-10%、表1)であった一方、メチルアルカンがn-アルカンに対して非常に少なく(図1)、カロテノイドが微量かつ散発的にしか含まれないことから、微生物マットが基礎生産性を支配せず76、浮遊性細菌とそれに対応するDOM分解産物が繁茂していたことが示唆された。 地層やバイオマーカー群の熱的成熟度が低いにもかかわらず、一般的に低い水素指数値(表1;特にレッドキノ試料)は、主に酸化的で浅海の縁辺環境における脂質の少ない有機物の投入と一致している。 また、微生物プランクトンが全体の基礎生産性や食物網の維持にどの程度寄与しているのかを明らかにすることはできない。 微生物マット群集はしばしば豊富な真核生物を含み、血栓症やクリンクリラミナイト相が顕著な南オマーンのアラグループ炭酸塩は、微生物マットの寄与が大きい新第三紀後期の環境の良い例であり、微細藻類の大きな寄与による豊富なステランシグナルをもたらした30,59。
24-IPCバイオマーカーのレベルが極めて低いことは、デモスッポンジが時に存在したもののこれらの環境では疎であったことを示唆している。 このことは、これらの低生産性環境では、エディアカラ生物相と競合する小型の濾過摂食動物の機会が少ないこと、波食帯より上部のエネルギーが高い浅海域の条件に対してデモスポンジの回復力が十分ではないこと、あるいは富栄養環境では光帯より下部で動的に維持される低酸素条件にスポンジがより適応していることを示すと思われる(図3)。 24-ipcはレッドキノ水平層のほとんどの試料で検出されなかったが、これらの試料は我々の試料群の中で一般にH/St比が最も高く、おそらくコトリン水平層の堆積時期までに生態系が変化したことを示していると思われる。 あるいは、海綿がこれらの環境に生息していたが、これらの診断用ステロイドバイオマーカーを大量に生産していなかったという可能性もあるが、24-ipc ステランはエディアカラ層の地層やオイルで最もよく検出される C30 ステラン化合物の一つで、我々の試料の一部でも検出されていることから、その可能性は低いと考えられる(Table 1)。
我々のデータで示唆された持続的な貧栄養海洋環境はエディアカラ海における局所的な条件であるが,先カンブリア時代の浅海域の海路では珍しいことではなかったと考えられる(図3). これまでのエディアカラ生物マーカーの研究の大部分は、富栄養環境で堆積した有機物に富む堆積岩とその石油製品に対して行われており、一般に微細藻類の寄与が大きい生物マーカー群が得られている44。 我々の結果は、海洋化学と生態系の場所による不均質性の規模をより正確に把握するために、適切な熱成熟度と異なる岩相を持つ有機物に富んだ地層を含む、より多様な堆積環境を研究することの重要性を強調している15, 3034, 59. ロディニア大陸の分裂期からエディアカラ紀にかけて、海洋換気が進み、化学風化や栄養供給が増加したにもかかわらず、海洋の不均質性は、栄養不足ではあるが、メタゾアの適応、競争、進化を育む居住環境を含む様々な海洋化学条件を地球海洋システムの中で維持した(図3)。 一方、富栄養化した海域では、南オマーン塩水盆(およびオマーン北部の厚いエディアカラ露頭)が顕著な例であり、浅く酸化的で生産性の低い大陸棚海域では、DOMやその他の有機デトリタスが局所的に利用可能であるにもかかわらず、逆にエディアカラ生物相の方が脱海綿動物よりも優先してコロニー化されるようになった。 しかし,エディアカラ海域の貧栄養環境が,巨視的多細胞生物の進化と海洋生態系に重要な役割を果たした可能性があることが明らかになった
。