Excepcionalmente baixa maturidade térmica dos estratos
A maturidade térmica é o fator mais importante que influencia a preservação dos biomarcadores sedimentares antigos, pois as moléculas orgânicas são suscetíveis a alterações estruturais e estereoquímicas durante o enterramento progressivo e com stress térmico crescente29. As rochas pré-cambrianas de maturidade térmica apropriada (maturidade da janela de óleo ou inferior) são pré-requisitos para a preservação de conjuntos de biomarcadores lipídicos robustos e primários. Múltiplos índices de estereoisômeros de marcadores de lúpano e esterano, juntamente com evidências independentes de baixos valores de Tmax (principalmente dentro de uma faixa de 417-433 °C, com uma média de 426 °C) da pirólise Rock-Eval, indicam que as rochas neste estudo representam de longe as rochas Ediacaran mais imaturas termicamente analisadas até o momento usando os métodos geoquímicos orgânicos de última geração (Tabela 1). Embora nossas rochas tenham sofrido diagênese sedimentar, elas não passaram significativamente para a janela de óleo, com exceção das amostras de 16PL ligeiramente mais maduras da bacia de Podillya na Moldávia, que ainda são adequadas para análise (maturação da janela de óleo de início a meio e sem sinais óbvios de contaminantes orgânicos). A maioria do conjunto então não tinha sido provavelmente exposta a temperaturas de enterramento superiores a 50 °C, pelo que as nossas amostras são altamente imaturas. Outras linhas de evidência molecular, que suportam baixa maturidade térmica e simgenicidade, incluem (i) domínio de alcanos policíclicos biomarcadores sobre n-alcanos em extratos de rochas (Fig. 2), (ii) sobrevivência de quantidades detectáveis de 17β,21β(H)-hopanos resolúvel a partir dos lúpanos mais abundantes e estáveis 17β,21α(H)- e 17α,21β(H)-configurações estereoquímicas (Tabela 1, Figs Suplementares. 1 e 2), (iii) uma preferência de n-alcanos ímpares entre os n-alcanos da faixa C22 a C27 (Fig. 2), uma vez que os n-alcanos mostram uma preferência de número de carbono somente antes da catagenese38, e (iv) a geração de marcadores de lúpano termicamente imaturos e esteranos a partir da fase de querogênio (insolúvel) usando hidropólise catalítica (Fig. 2 Suplementar). A baixa maturidade térmica da matéria orgânica sedimentar em nossas amostras é consistente com indicadores de alteração sedimentar previamente publicados, incluindo índices de alteração de conodont e acritarch, e estudos anteriores de pirólise Rock-Eval e biomarcadores das rochas sedimentares Ediacaran e Paleozoic em Baltica36,39,40, bem como mineralogia argilosa41. Criticamente, em ambientes onde a maturidade térmica é tão baixa, os mecanismos para alteração significativa dos sinais primários δ15N e δ13C estão em grande parte ausentes42,

Distribuição de hidrocarbonetos alifáticos extraíveis para uma amostra representativa. um Cromatograma iônico total (TIC) para hidrocarbonetos alifáticos extraíveis para Lugovoe #13-73 m do Horizonte Redkino. As séries n-alcanos, pristano (Pr), fitano (Ph) e C27-C34 hopanes (denotados pelo seu número total de carbono e estereoquímica em C-17, C-21 e C-22, por exemplo, C31αβR) são rotulados; ** denota C29 e C30 hopenes. Note a dominância da abundância de lúpano sobre outras classes de compostos alcanos. b Cromatograma parcial de íons 85-Da mostra dominância da série n-alcanos, exibindo uma preferência de número ímpar ou ímpar de carbono na faixa C22-C27, sobre metilalcanos
Padrão de montagem de biomarcadores lípidos
Uma relação de abundância dos lúpanos principais (C27-C35) para os esteranos principais (C27-C29) é frequentemente usada para avaliar o equilíbrio das entradas de organismos de fonte bacteriana versus eucariótica no antigo ecossistema aquático. Hopanos são fósseis moleculares derivados de hopanoides, que são lípidos de membrana celular sintetizados por uma grande variedade de grupos bacterianos. Da mesma forma, os esteranos são derivados de precursores de esterol, que são produzidos quase exclusivamente por eucariotas43. As proporções de Hopano/sterano (H/St) para as nossas amostras cobrem uma faixa de valores surpreendentemente grande de 1,6 a 119,2 (Tabela 1). Para o contexto, as razões H/St de rochas e óleos neoproterozóicos ricos em orgânicos caem normalmente numa faixa estreita de 0,5 a 2.030,31,32,33. Embora exista uma clara diferença entre os valores do jovem Horizonte Kotlin (média H/St de 8,9) e do antigo Horizonte Redkino (média H/St de 42,9), os valores para a maioria das amostras são anormalmente elevados e sugerem contribuições anormalmente elevadas de bactérias.
Por último o Neoproterozóico, as algas eucarióticas foram um componente ecologicamente significativo e os principais produtores em muitos ecossistemas marinhos15,34,44. A discrepância entre a contribuição globalmente importante das algas eucarióticas no Neoproterozóico tardio e ainda os baixos níveis de biomarcadores esteranos nestas amostras indica que deve haver algum determinante local na abundância eucariótica. A explicação mais parcimoniosa para as proporções extremamente elevadas de lúpano/sterano ao lado de baixos teores de carbono orgânico total (COT) e baixos índices de hidrogênio (IH) encontrados para nossas amostras (Tabela 1) é que esses estratos foram depositados em ambientes oligotróficos (ou seja, fortemente limitados em nutrientes), nos quais as bactérias superaram as algas combatidas. Enquanto os analógicos modernos para os antigos mares epicratônicos desenvolvidos durante o alto nível do mar são difíceis de encontrar, observações paralelas de proporções mais elevadas de lúpano/sterano (por ordem de magnitude) foram encontradas anteriormente para rochas sedimentares orgânicas limpas versus rochas sedimentares ricas em orgânicos, depositadas em mares epicontinentais ordoviciano-silurianos45.
Tipicamente, baixo teor de COT pode ser causado pela deposição limitada de matéria orgânica em ambientes de baixa produtividade, baixo potencial de conservação de matéria orgânica na coluna de água ou sedimentos, ou diluição com alto fluxo siliciclástico. Nos oceanos modernos, a produtividade é mais comumente limitada por baixos níveis dos nutrientes essenciais: nitrogênio, fósforo, silício e ferro46. Nitrogênio, fósforo e ferro são limitados em regiões a céu aberto onde o afloramento, a entrada de poeira e o escoamento da costa não fornecem quantidades suficientes de nutrientes. Em ambientes oligotróficos modernos, a razão entre a biomassa bacteriana e a eucariótica é maior do que em ambientes eutróficos ou mesotróficos. Curiosamente, as amostras de Podillya 16PL do horizonte Redkino da Moldávia são lodos contendo fósforo e produzem proporções significativamente mais baixas de lodo/sterano (8,1:11,5) do que as outras amostras Redkino da Tabela 1 (embora ainda significativamente mais altas do que as encontradas na Bacia Salina de Omã do Sul ou em sedimentos Fanerozóicos ricos em fósforo orgânico, que tipicamente se situam dentro de um estreito, 0.5-2,0), possivelmente sugerindo que uma maior disponibilidade de fosfatos poderia ter influenciado favoravelmente a razão eucariótica/bacteriana encontrada localmente (dado que as assinaturas isotópicas de nitrogênio são amplamente invariantes, veja a próxima seção). O conteúdo de fósforo (P), assim como as relações P/Fetotal e P/Al, são geralmente baixos para os sedimentos Ediacaran de Podillya, Ucrânia e Estónia47; com excepção deste nível estratigráfico marcado com nódulos de fosforito. Similarmente, níveis baixos (~0,01 a ~0,1 wt%) de P em rochas siliciclásticas de Kotlin e Redkino foram relatados a partir de um núcleo de perfuração da margem nordeste da Plataforma do Leste Europeu11.
A topografia ampla e rasa nos mares epicontinentais através do Báltico poderia ter sustentado fósforo ou outras limitações nutricionais em cenários marginais, devido à precipitação autógena de fosfato com minerais de ferro nas águas de superfície óxicas e seqüestro de uma gama de elementos por sedimentos de prateleira48. Se o fosfato fosse um nutriente limitante, o aumento do fósforo biodisponível teria aumentado a produção primária local e proporcionado condições de crescimento mais favoráveis para o fitoplâncton unicelular de maior tamanho49,50,51. Com respeito ao moderno sistema oceânico, as picocianobactérias marinhas, Prochlorococcus e Synechococcus, são reconhecidas por dominarem as contagens de células fitoplâncton e a biomassa nos ambientes oceânicos oligotróficos tropicais e subtropicais, incluindo as regiões oligotróficas limitadas por fosfato das águas de superfície oceânica52. O proclorococo e o SAR11 heterotrófico (Pelagibacter) florescem devido a uma variedade de adaptações, incluindo baixos custos de energia em virtude de pequenos genomas e baixas taxas de replicação, uma maior relação superfície-volume através de menores tamanhos de células e funções adicionais de absorção de células para maximizar a utilização de nutrientes53. Muitas bactérias também são capazes de substituir nutrientes de baixa abundância, por exemplo, utilizando sulfolipídios em vez de lipídios com fósforo em ambientes com déficit de P54, ou pelo uso de substratos alternativos, por exemplo, obtendo nitrogênio da atmosfera N2 através da fixação de nitrogênio55, para aliviar o estresse por nutrientes. Um estudo recente sobre a sobrevivência do bacterioplâncton marinho em ambientes oligotróficos56 , onde o fosfato disponível é limitado, sugere um papel importante para o metabolismo dos polifosfatos nos oligotróficos marinhos. Os eucariotas selecionados que competem em ambientes oligotróficos são tipicamente pequenos picoeukaryotes, que podem complementar suas necessidades nutricionais através da mixotrofia57. A razão biomassa do picoeukaryote para cianobactérias tende a aumentar sob o aumento do fornecimento de nutrientes49. A possível influência do fósforo, e de outros nutrientes biolimitantes, na moderação da produtividade primária e da estrutura da comunidade marinha nos mares epeíricos de Ediacaran requer mais investigação.
Sem exceção, a abundância de esteranos C29 é maior do que os correspondentes esteranos C27 ou C28 para todos os nossos locais (Tabela 1). A predominância de esteranos C29 sobre C27 e C28 indica provavelmente uma dominância de algas verdes dentro da comunidade fitoplanctônica eucariótica44,58. Esta característica tem sido observada na maioria dos estudos anteriores de biomarcadores Ediacaran15,30,31,32,33,34,35,36,44. Notavelmente, a distribuição de esterano C30 em várias amostras de cada núcleo de perfuração no Horizonte de Kotlin contém quantidades baixas, mas detectáveis, do biomarcador de demosponge sterane conhecido como 24-isopropilcolestano (24-ipc)59,60. No total, 24-ipc steranes foram relatados em amostras de rochas e óleo datando de tão longe quanto o criogênico (>635 Ma atrás) na Bacia Salina Sul de Omã e representam a evidência mais antiga do biomarcador lipídico para metazoans59,60. Os biomarcadores 24-ipc em nossas amostras eram cerca de uma ordem de magnitude menor em abundância (em relação ao total de C27 a C30, estas foram apenas 0,06-0,61%; média = 0,22%) em comparação com as rochas e óleos Ediacaran do Sul de Omã (1,7% em média59) ou estavam abaixo dos limites de detecção devido à abundância negligenciável para a maioria das amostras.
Nitrogênio e isótopos orgânicos de carbono
Isótopos de nitrogênio podem ajudar a discernir o equilíbrio relativo no ciclo do nitrogênio, e o grau em que a fixação de nitrogênio ou a desnitrificação incompleta foram caminhos significativos para influenciar o equilíbrio de nutrientes disponíveis para as comunidades marinhas. Quando as bactérias diazotróficas fixam nitrogénio molecular devido à falta de nitrogénio fixo na coluna de água, isto pode produzir valores de isótopos sedimentares de nitrogénio a granel próximos de 0‰61. Isto está em contraste com as assinaturas positivas de isótopos de nitrogênio (na faixa de +2 a +10‰) com um modo de +4 a +6‰ encontrado para os sedimentos marinhos neoproterozóicos depositados sob o que se pensa serem condições nitrato-repletas, onde o nitrato sofreu apenas desnitrificação parcial62. Os valores de isótopos de nitrogênio para todas as nossas amostras bar um outlier (Tabela 1) cobrem uma variação positiva limitada de +3,5 a +6,5‰, que se sobrepõe ao modo para rochas sedimentares Neoproterozóicas tardias, ricas em matéria orgânica62. Os nossos dados indicam que o nitrato dominava a piscina de nitrogênio inorgânico dissolvido, sendo que as condições da coluna d’água óxica prevaleceram. A faixa de valores de δ15N também sugere que a tributação N2- não foi o modo primário de aquisição de nitrogênio para produtores primários. Ao contrário, o ciclo do nitrogênio foi provavelmente dominado pela reciclagem da coluna d’água, e o N orgânico foi sujeito à oxidação quantitativa para o nitrato, como acontece nos ambientes marinhos modernos e próximos. A desnitrificação bentônica e o enterramento de N orgânico teriam sido os sumidouros primários de nitrogênio inorgânico dissolvido, e a faixa de valores de δ15N sugere apenas um papel limitado para a desnitrificação incompleta da coluna d’água, o que tipicamente resulta em um significativo enriquecimento de 15N63. A variabilidade limitada nos valores de δ15N, portanto, provavelmente reflete δ15N de nitrato aventado na plataforma, com pequena influência da limitada redução de nitrato na coluna d’água e fixação de N2. Sem forte evidência de fixação de nitrogênio no δ15N, nós supomos que o nitrogênio não era o nutriente primário biolimitante nas bacias epicontinentais de Baltica. O isolamento de fontes fluviais e eólicas de fósforo, e as condições óxicas nas bacias epicontinentais amplas e pouco profundas podem ter aumentado a remoção do fósforo autógeno e das fases que contêm traços metálicos, resultando em limitação de nutrientes que restringiram o crescimento e a produção de células eucarióticas.
Os valores dos isótopos de carbono orgânico total (CTOC) variam de -23,0 a -33,9‰, com as maiores diferenças observadas entre as diferentes localizações dos núcleos de perfuração. As proporções relativas de 13C-enriquecimento para isótopos superiores a ca. -28‰ contrastam com os dados dos estratos contemporâneos depositados nas configurações eutróficas de Oman30,31. Entretanto, o intervalo que relatamos é geralmente consistente com o intervalo δ13CTOC para outros locais do Baltica depositados no mesmo período de tempo11. A diferença entre Baltica e Omã pode destacar o fato de que os valores de δ13CTOC não refletem exclusivamente uma mudança secular uniforme no ciclo do carbono durante esse intervalo de tempo31,64. Ao contrário, a faixa δ13CTOC de Baltica pode, em parte, refletir a ecologia microbiana dominada bacterialmente sugerida pelas proporções únicas de biomarcadores lipídicos. O tamanho das células pequenas, as relações superfície/volume elevadas e as taxas de crescimento lento sob condições oligotróficas podem aumentar a magnitude do fracionamento durante a autotrofia (εp)65,66, resultando em baixos valores δ13C.
O mecanismo para as assinaturas isotópicas mais 13C-enriquecidas dentro da faixa relatada11 para a sucessão tardia de Ediacaran Baltica é menos claro. A contribuição potencial da matéria orgânica detrital, metamorficamente alterada, para as sucessões sedimentares pré-cambrianas de baixo TOC é um possível mecanismo para as disparidades entre os sites em δ13CTOC64. No entanto, o conteúdo de TOC das nossas amostras, embora baixo, é geralmente superior a 0,10 wt%, não revelando qualquer relação entre o conteúdo de TOC e os valores de δ13C. Além disso, as nossas amostras contêm matéria orgânica termicamente imatura e padrões lipídicos biomarcadores que são inconsistentes com uma fonte de carbono principalmente alóctone. Mais 13C-enriquecidas δ13CTOC assinaturas podem ser o resultado de mecanismos alternativos para a assimilação de carbono. Mecanismos de concentração de carbono ou absorção ativa de bicarbonato por procariotas pode resultar em valores menores para εp e maiores para δ13CTOC valores67,68. O surgimento da biota Ediacaran pode ter expandido significativamente as teias alimentares marinhas e estimulado novas vias de heterotrofia microbiana, incluindo possíveis contribuições de ciclos complexos de carbono dentro de esteiras microbianas bentônicas. Com uma consideração adicional pelo papel potencialmente importante da matéria orgânica dissolvida69 como fonte de carbono, a amplitude dos valores de δ13CTOC provavelmente reflecte uma gama de processos biogeoquímicos de reciclagem de carbono que podem ser únicos no ambiente marinho de Ediacaran em evolução e podem estar relacionados com um fenómeno para o qual não temos análogos modernos representativos.
Assemblagens de biomarcadores lipídicos antigos e indicadores isotópicos estáveis para a produção de esteiras bentónicas e microbianas em tandem com limitação fixa de azoto foram encontrados nas primeiras rochas triássicas do Sul da China70. No rescaldo da extinção da massa permiana final em Meishan, houve um grande pico na proporção hopane/sterane (até cerca de 60) acompanhado por uma forte mudança na assinatura isotópica de Norg para valores de 0 a -2‰, consistente com a diazotrofia bacteriana. Isto está associado a um alto sinal de 2-metilhopanos (índice de 2-metilhopanos até 33%) e metilalcanos característicos que apontam para a proliferação de esteiras microbianas bentônicas. Não observamos uma tendência similar na assinatura de isótopos de nitrogênio ou padrões de biomarcadores que obviamente apontariam para entrada significativa de esteiras microbianas em nosso conjunto de dados Ediacaran, embora um valor singular de 15N tenha sido encontrado em nossas amostras (Tabela 1). Portanto, o forte sinal bacteriano observado em nossas amostras de Baltica provavelmente não é predominantemente uma assinatura de esteiras microbianas bentônicas.
Paleoenvironmental sustenance of Ediacara biota vs. demosponges
As condições oligotróficas aparentes através das bacias epicratônicas e da margem continental do Báltica, uma vez que ele derivou de latitudes altas para baixas do Ediacaran tardio para o Cambriano precoce, podem estar associadas ou com o advento limitado de águas profundas relativamente enriquecidas com nutrientes ou com o esgotamento de nutrientes, resultantes da assimilação e da depleção durante o transporte e deposição através dessas bacias epicontinentais amplas e rasas-marinhas (Fig. 3). Estas bacias foram episodicamente isoladas dos oceanos e desenvolveram hipersalinidade (por exemplo, durante o tempo Redkino) e condições salobras (por exemplo, durante o tempo Kotlin71). A estabilidade tectônica de longo prazo resultou em uma topografia de baixo relevo do Ediacaran Baltica tardio, altamente suscetível a inundações e suprimento ineficiente de fósforo derivado de intempéries. Em contraste ao Baltica, estudos biomarcadores dos estratos do Supergrupo Huqf em Omã revelaram um ecossistema eutrófico, rico em microalgas30,31,59, mas sem a biota Ediacara, mesmo em extensos afloramentos nas montanhas de Omã e na região de Huqf, em cenários de prateleira interna a externa3. A paleogeografia da Bacia Salina do Sul de Omã foi reconstruída para o Neoproterozóico tardio a ~13° do equador no hemisfério sul, amplamente semelhante à paleolatitude do Baltica72.
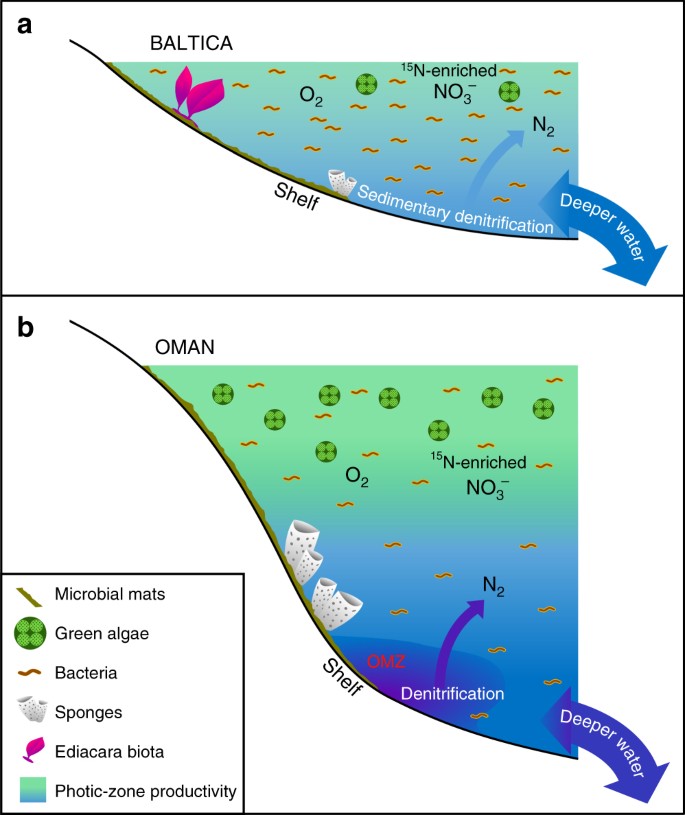
Melhores diferenças em ambientes marinhos Ediacaran de baixa produtividade vs. produtiva. Diagramas esquemáticos são mostrados para uma extensa bacia epicontinental oligotrófica e marinha rasa do Baltica frequentemente dominada pela produtividade bacteriana onde a fauna de corpo mole da Ediacara floresceu e onde a desnitrificação e anammox provavelmente foram restritas a sedimentos; e b eutróficas e marinhas mais profundas da Bacia do Sal de Omã do Sul, onde as algas verdes prosperavam como produtoras primárias e as demosponges eram abundantes, mas a fauna de corpo mole de Ediacara não era proeminente (com os fósseis biota de Ediacara também ausentes nos afloramentos correlativos de Ediacaran no norte de Omã). Nas margens continentais produtivas, desnitrificação e anammox provavelmente ocorreram tanto na coluna d’água quanto nos sedimentos
Não há evidências de anóxia persistente e extensa nas margens continentais rasas do Baltica durante o Ediacaran tardio11, e sugerimos que as condições oligotróficas aqui descritas foram causadas por fluxos ineficientes de nutrientes terrestres e de águas profundas para esses ambientes, amplamente semelhantes aos de ambientes oligotróficos nos sistemas oceânicos modernos. De facto, uma investigação geoquímica de elementos vestigiais do Utkina Zavod e núcleos de perfuração adjacentes da área de São Petersburgo sugere que as nossas amostras foram depositadas sob condições óxicas73. Esta interpretação é suportada independentemente pelos nossos dados das distribuições estendidas de lúpano (C31-C35) que caem acentuadamente em abundância acima dos compostos C31 com número crescente de carbono, o que é característico da degradação da cadeia lateral dos bacteriohopanepolióis durante a diagênese sob condições óxicas29, e também por índices baixos de hidrogênio, como medido pela pirólise Rock-Eval (todos inferiores a 230 mg/g COT, e muitos abaixo de 100 mg/g COT) para estas amostras imaturas devido à formação de querogênio recalcitrante via degradação oxidativa e recondensação de biomassa primária pobre em lipídios em ambientes localmente óxicos (Tabela 1). Os organismos multicelulares Ediacaran que existiam nestes ambientes marinhos rasos devem ter tido substratos orgânicos suficientes para a heterotrofia para satisfazer as suas necessidades alimentares, juntamente com oxigénio e outros nutrientes suficientes para sustentar o seu metabolismo. As bacias epicontinentais do Báltica eram provavelmente mais persistentemente óxicas do que os ambientes altamente produtivos que margeavam as zonas de oxigénio-mínimo, como no meio da plataforma externa da Bacia do Sal de Omã do Sul, onde a respiração da biomassa planctónica abundante teria mantido o oxigénio dissolvido mais baixo abaixo da zona fótica.
Ediacara biota e outros organismos multicelulares que vivem no mar do Báltico teriam que lidar com a mudança das fontes alimentares (bacterianas vs. eucarióticas), uma vez que os fluxos de nutrientes variaram ao longo do tempo, incluindo pequenas células e detritos orgânicos em bacias epicontinentais onde as bactérias eram os produtores primários dominantes (Fig. 3). As comunidades modernas de recife de coral sobrevivem em ambientes oligotróficos tropicais devido à reciclagem eficiente dos nutrientes, incluindo a geração de um fluxo de matéria orgânica dissolvida (DOM) que ajuda a manter a heterotrofia faunística dentro do ecossistema do recife74. O estabelecimento de uma estrutura trófica marinha com organismos eucarióticos multicelulares sustentados pela alimentação com detritos orgânicos teve que pós-datar a expansão ambiental à escala global dos eucariotas em diversos ambientes marinhos, que ocorreu através do intervalo Toniano-Criogénico (ca. 800-635 Ma), conforme aferido a partir de registos biomarcadores15. Poderíamos então esperar discernir evidências de diferenças significativas na comunidade marinha e estrutura trófica da localidade para a localidade durante o final do período Ediacaran, com o equilíbrio de nutrientes local selecionando comunidades microbianas ricas em eucariotas ou eucariotas-lean e com a expansão progressiva de organismos multicelulares adicionando outra dimensão de complexidade a nível de organismo e comunidade. Um contraste regional significativo na disponibilidade de fosfatos e outros nutrientes em ambientes de prateleira é também uma consequência esperada de uma estrutura redox marinha global heterogênea para o Ediacaran tardio, antes da oxigenação do oceano profundo38.
A dominância da produtividade bacteriana do picoplâncton e produtos de degradação da matéria orgânica dissolvida (DOM) associada nas bacias epicontinentais do Baltica poderia ter sustentado um ecossistema de laço microbiano em paralelo com a estrutura trófica convencional baseada em cerca de produtores planctônicos de maior tamanho50. Isto pode ter favorecido diferentes modos de heterotrofia, incluindo a alimentação em suspensão e, possivelmente, a osmotrofia, como estratégia de alimentação viável para alguns rangeomorfos, esponjas e outros organismos multicelulares Ediacaran tardios18 em conjunto com a heterotrofia ativa (motil) emergente19,20. Os sistemas aquáticos limitados em nutrientes, como regra geral, são frequentemente dominados por pequenos fitoplâncton unicelulares e plâncton heterotrófico, com bactérias que superam os eucariotas, e sustentando biomassa líquida baixa em ambientes marinhos oligotróficos dos oceanos modernos49,51. O DOM é uma importante fonte de nutrientes orgânicos e muitas vezes controla a produtividade e a biomassa líquida nos mares tropicais oligotróficos modernos, mas também o é a co-limitação de nitrogênio e fósforo75. Da mesma forma, o DOM também era provavelmente um substrato importante para sustentar bactérias heterotróficas e um laço microbiano em ambientes oligotróficos antigos. As esteiras microbianas bentónicas podem ter sido um componente desta teia alimentar dominada por bactérias e os solos das esteiras têm sido implicados para melhorar a preservação da Ediacara biota em ambientes marinhos7. Enquanto valores moderados de índice 2-metilhopano foram encontrados para um subconjunto de nossas amostras (4-10%, Tabela 1), quantidades muito baixas de metilalcanos em relação aos n-alcanos (Fig. 1), e apenas vestígios e ocorrências esporádicas de carotenóides sugerem que as esteiras microbianas não dominaram a produtividade primária76, e que as bactérias planctônicas e, correspondentemente, seus produtos de decomposição DOM floresceram. Os valores geralmente baixos do índice de hidrogênio (Tabela 1; especialmente para amostras Redkino), apesar da baixa maturidade térmica dos estratos e dos biomarcadores, também são consistentes com a entrada orgânica pobre em lipídios, principalmente em ambientes marginais marinhos rasos e óxicos. As ocorrências de matrizes microbianas, aferidas apenas a partir de texturas sedimentológicas, não nos dizem respeito ao equilíbrio de eucariotas (microalgas) com bactérias em qualquer caso, nem podem limitar a contribuição relativa do plâncton microbiano para a produtividade primária global e a sustentação das teias alimentares. As comunidades de esteiras microbianas que frequentemente contêm eukaryotes abundantes e os carbonatos do Grupo Ara de South Oman com proeminentes trombolíticos e fácies laminitas enrugadas são um bom exemplo de ambientes neoproterozóicos tardios com contribuição significativa de esteiras microbianas, produzindo abundantes sinais esteranos devido a uma grande contribuição de microalgas30,59.
O nível extremamente baixo de biomarcadores 24-ipc sugere que, embora as demosponjas estivessem por vezes presentes, elas eram esparsas nestes ambientes. Isto poderia indicar menos oportunidades para animais de alimentação filtrada menores em competição com a biota Ediacara nestes ambientes de baixa produtividade, resiliência insuficiente das demosponges contra condições marinhas rasas mais energéticas acima da base da onda de tempo justo, ou melhor adaptabilidade das esponjas a condições de baixa oxigenação mantida dinamicamente abaixo da zona fótica em ambientes eutróficos (Fig. 3). No total, 24-ipc não foi detectado na maioria das amostras do Horizonte Redkino, que também têm geralmente as maiores relações H/St entre o nosso conjunto de amostras, talvez indicando uma mudança ecológica pelo tempo de deposição do Horizonte Kotlin. Alternativamente, também é plausível que as esponjas habitaram estes cenários, mas não produziram estes biomarcadores esteróides de diagnóstico em abundância, embora isto pareça menos provável, uma vez que o esterano C30 está entre os compostos C30 mais frequentemente detectados nos estratos e óleos Ediacaran30,3133,59,60 e também é detectável num subconjunto das nossas amostras (Tabela 1).
Embora os ambientes marinhos oligotróficos persistentes sugeridos por nossos dados representem condições localizadas nos oceanos de Ediacaran, eles provavelmente não foram incomuns para os mares pré – cambrianos rasos-marinhos (Fig. 3). A grande maioria dos estudos biomarcadores Ediacaran anteriores foram conduzidos em rochas sedimentares ricas em matéria orgânica depositadas em cenários eutróficos e seus produtos petrolíferos, que geralmente produzem conjuntos biomarcadores consistentes com significativa contribuição de fontes microalgas44. Nossos resultados destacam a importância do estudo de uma maior variedade de ambientes deposicionais, incluindo estratos organo-matéria limpos de maturidade térmica apropriada e diferentes litologias15.3034.59, a fim de obter uma imagem mais precisa da escala de heterogeneidade da química marinha e da ecologia de local para local. Apesar da progressiva ventilação oceânica e do aumento da meteorologia química e do fornecimento de nutrientes durante a quebra da Rodínia e durante todo o período Ediacaran77,78, a heterogeneidade oceânica manteve uma variedade de condições químicas marinhas, incluindo ambientes pobres em nutrientes, mas habitáveis, que promoveram a adaptação, competição e evolução dos metazoários dentro do sistema oceânico global (Fig. 3). Enquanto as prateleiras marinhas eutróficas hospedavam demospongas, mas muitas vezes faltava o Ediacara biota, sendo a Bacia do Sal de Omã do Sul (e os afloramentos de Ediacaran espessos correlativos mais ao norte, em Omã) um exemplo proeminente; contraintuitivamente, os mares rasos, óxicos e epicontinentais menos produtivos foram colonizados pelo Ediacara biota, em preferência às demospongas, apesar do DOM e outros detritos orgânicos estarem disponíveis localmente para alimentação. Se os requisitos metabólicos ou a pressão seletiva ambiental restringiram a biota multicelular de corpo mole do Ediacaran a esses ambientes permanece incerta; entretanto, nosso estudo destaca que os ambientes oligotróficos Ediacaran desempenharam um papel potencialmente crucial na evolução dos organismos multicelulares macroscópicos e da ecologia da comunidade marinha.