Výjimečně nízká teplotní zralost vrstev
Termální zralost je nejdůležitějším faktorem ovlivňujícím zachování starobylých sedimentárních biomarkerů, protože organické molekuly jsou náchylné ke strukturním a stereochemickým změnám během postupného pohřbívání a s rostoucím tepelným namáháním29. Prekambrické horniny vhodné tepelné zralosti (zralost ropného okna nebo nižší) jsou předpokladem pro zachování robustních a primárních biomarkerových lipidových souborů. Mnohonásobné poměry stereoizomerů biomarkerů hopanu a steranu spolu s nezávislými důkazy z nízkých hodnot Tmax (většinou v rozmezí 417-433 °C, s průměrem 426 °C) z pyrolýzy Rock-Eval naznačují, že horniny v této studii představují zdaleka nejvíce tepelně nezralé ediakarské horniny, které byly dosud analyzovány pomocí nejmodernějších organických geochemických metod (tab. 1). Přestože naše horniny prošly sedimentární diagenezí, nepřešly výrazně do ropného okna, kromě poněkud zralejších vzorků z výchozu 16PL z Podolské pánve v Moldavsku, které jsou stále vhodné k analýze (raná až střední zralost ropného okna a bez zjevných známek organických kontaminantů). Většina souboru pak pravděpodobně nebyla vystavena pohřebním teplotám přesahujícím 50 °C, naše vzorky jsou tedy velmi nezralé. Mezi další molekulární důkazy, které podporují nízkou tepelnou zralost a syngenicitu, patří (i) převaha polycyklických biomarkerových alkanů nad n-alkany v horninových extraktech (obr. 2), (ii) přežívání detekovatelného množství 17β,21β(H)-hopanů rozlišitelných od hojnějších hopanů disponujících stabilními 17β,21α(H)- a 17α,21β(H)-stereochemickými konfiguracemi (tab. 1, doplňkové obr. 1 a 2). 1 a 2), (iii) zřetelná preference lichých nad sudými n-alkany v rozmezí C22 až C27 (obr. 2), neboť n-alkany vykazují preferenci počtu uhlíků pouze před katagenezí38, a (iv) generace tepelně nezralých hopanových a steranových biomarkerů z (nerozpustné) kerogenní fáze pomocí katalytické hydropyrolýzy (doplňkový obr. 2). Nízká termická zralost sedimentární organické hmoty v našich vzorcích je v souladu s dříve publikovanými indikátory sedimentární alterace, včetně indexů konodontní a akritarchové alterace, a s předchozími studiemi pyrolýzy a biomarkerů Rock-Eval ediakarských a paleozoických sedimentárních hornin v Baltice36,39,40 a také mineralogie jílů41. Kritické je, že v prostředí, kde je tepelná zralost takto nízká, mechanismy pro významnou alteraci primárních signálů δ15N a δ13C do značné míry chybí42.

Rozdělení extrahovatelných alifatických uhlovodíků pro reprezentativní vzorek. a Celkový iontový chromatogram (TIC) pro extrahovatelné alifatické uhlovodíky pro Lugovoe #13-73 m z Redkinského horizontu. Označeny jsou n-alkanové řady, pristan (Pr), fytan (Ph) a hopeny C27-C34 (označené podle celkového počtu uhlíků a stereochemie na C-17, C-21 a C-22, např. C31αβR); ** označuje hopeny C29 a C30. Všimněte si převahy množství hopenů nad ostatními třídami alkanických sloučenin. b Částečný iontový chromatogram 85-Da ukazuje dominanci n-alkanové řady, která vykazuje zřetelnou preferenci lichého nad sudým počtem uhlíků v rozsahu C22-C27, nad methylalkany
Soubory lipidových biomarkerů
Poměr abundance hlavních (C27-C35) hopanů a hlavních (C27-C29) steranů se často používá k posouzení rovnováhy mezi vstupy bakteriálních a eukaryotických zdrojových organismů do starého vodního ekosystému. Hopany jsou molekulární fosilie odvozené od hopanoidů, což jsou lipidy buněčných membrán syntetizované nejrůznějšími skupinami bakterií. Podobně sterany jsou odvozeny od prekurzorů sterolů, které jsou produkovány téměř výhradně eukaryoty43. Poměry hopanů a steranů (H/St) u našich vzorků pokrývají nápadně velký rozsah hodnot od 1,6 do 119,2 (tab. 1). Pro kontext, poměry H/St z neoproterozoických hornin a olejů bohatých na organické látky obvykle spadají do úzkého rozmezí od 0,5 do 2,030,31,32,33 . I když je zřejmý rozdíl v hodnotách z mladšího kotlinského horizontu (průměrná hodnota H/St 8,9) a staršího redkinského horizontu (průměrná hodnota H/St 42,9), hodnoty pro většinu vzorků jsou neobvykle vysoké a naznačují anomálně zvýšený příspěvek bakterií.
Koncem neoproterozoika byly eukaryotické řasy ekologicky významnou složkou a hlavními producenty v mnoha mořských ekosystémech15,34,44 . Rozpor mezi globálně významným příspěvkem eukaryotických řas v pozdním neoproterozoiku a přesto nízkými hladinami biomarkerů steranu v těchto vzorcích naznačuje, že musí existovat nějaký lokální determinant na početnost eukaryot. Nejjednodušším vysvětlením extrémně zvýšených poměrů hopanu a steranu spolu s nízkými obsahy celkového organického uhlíku (TOC) a nízkými vodíkovými indexy (HI) zjištěnými u našich vzorků (tab. 1) je, že tyto vrstvy byly uloženy v oligotrofním (tj. živinami silně omezeném) prostředí, v němž bakterie konkurovaly řasám. Zatímco moderní analogy pro starobylá epikratonická moře vzniklá během vysokého stání mořské hladiny se hledají obtížně, paralelní pozorování vyšších poměrů hopanu a steranu (až o řád) byla již dříve zjištěna pro organicky chudé oproti organicky bohatým sedimentárním horninám usazeným v ordovicko-silurských epikontinentálních mořích45.
Typicky může být nízký obsah TOC způsoben omezenou depozicí organické hmoty v prostředí s nízkou produktivitou, nízkým konzervačním potenciálem organické hmoty ve vodním sloupci nebo sedimentech nebo zředěním vysokým tokem siliciklastik. V moderních oceánech je produktivita nejčastěji omezena nízkým obsahem základních živin: dusíku, fosforu, křemíku a železa46. Dusík, fosfor a železo jsou omezeny v oblastech otevřeného oceánu, kde upwelling, vstup prachu a pobřežní splachy nedodávají dostatečné množství živin. V moderním oligotrofním prostředí je poměr bakteriální a eukaryotické biomasy vyšší než v eutrofním nebo mezotrofním prostředí. Zajímavé je, že vzorky z výchozu Podillya 16PL z redkinského horizontu v Moldavsku jsou slínovce obsahující fosforit a poskytují výrazně nižší poměr hopanů a steranů (8,1:11,5) než ostatní redkinské vzorky v tabulce 1 (i když stále výrazně vyšší než ty, které byly nalezeny v jihoománské solné pánvi nebo v organicky bohatých fanerozoických sedimentech, které obvykle spadají do úzkého, 0.5-2,0), což možná naznačuje, že zvýšená dostupnost fosfátů mohla příznivě ovlivnit lokálně zjištěný poměr eukaryot k bakteriím (vzhledem k tomu, že izotopové znaky dusíku jsou do značné míry neměnné, viz následující oddíl). Obsahy fosforu (P), stejně jako poměry P/Fetotal a P/Al, jsou jinak pro pozdně ediakarské sedimenty Podolí, Ukrajiny a Estonska47 obecně nízké; s výjimkou této stratigrafické úrovně vyznačené fosforovými konkrecemi. Podobně nízké (~0,01 až ~0,1 hm. %) obsahy P v kotlinských a redkinských siliciklastických horninách byly zaznamenány z vrtného jádra ze severovýchodního okraje Východoevropské platformy11.
Široká a mělká topografie v epikontinentálních mořích napříč Baltikou mohla udržovat omezení fosforu nebo jiných živin v okrajových polohách v důsledku autogenního srážení fosfátů s minerály železa v oxických povrchových vodách a sekvestrace řady prvků šelfovými sedimenty48. Pokud byl fosfát limitující živinou, zvýšení biologicky dostupného fosforu by zvýšilo místní primární produkci a poskytlo příznivější podmínky pro růst jednobuněčného fytoplanktonu větších rozměrů49,50,51. Pokud jde o současný oceánský systém, je známo, že mořské picocyanobakterie, Prochlorococcus a Synechococcus, dominují počtu buněk a biomase fytoplanktonu v oligotrofním prostředí tropických a subtropických oceánů, včetně oligotrofních oblastí povrchových vod oceánů omezených fosfáty52. Prochlorococcus a heterotrofní SAR11 (Pelagibacter) prosperují díky řadě adaptací, včetně nízkých energetických nákladů díky malým genomům a nízké rychlosti replikace, vyššímu poměru plochy k objemu díky menším rozměrům buněk a dalším funkcím buněčného příjmu pro maximální využití živin53. Mnoho bakterií je také schopno nahradit živiny s nízkým obsahem, např. využitím sulfolipidů namísto lipidů obsahujících fosfor v prostředí s nedostatkem P54 , nebo využitím alternativních substrátů, např. získáváním dusíku z atmosférického N2 prostřednictvím fixace dusíku55 , aby zmírnily nutriční stres. Nedávná studie týkající se přežívání mořského bakterioplanktonu v oligotrofním prostředí56 , kde je dostupný fosfát omezený, naznačuje důležitou roli metabolismu polyfosfátů u mořských oligotrofů. Vybraná eukaryota, která soutěží v oligotrofním prostředí, jsou obvykle malá picoeukaryota, která mohou doplňovat své nutriční požadavky prostřednictvím mixotrofie57. Poměr biomasy picoeukaryot a cianobakterií má tendenci se při zvýšeném přísunu živin zvyšovat49. Možný vliv fosforu a dalších biolimitujících živin na mírnění primární produktivity a struktury mořských společenstev v ediakarských epeirických mořích vyžaduje další zkoumání.
Bez výjimky je na všech našich lokalitách hojnost steranů C29 vyšší než odpovídajících steranů C27 nebo C28 (tab. 1). Převaha steranů C29 nad sterany C27 a C28 pravděpodobně ukazuje na dominanci zelených řas ve společenstvu eukaryotického fytoplanktonu44,58. Tento rys byl pozorován ve většině předchozích ediakarských studií biomarkerů15,30,31,32,33,34,35,36,44. Pozoruhodné je, že distribuce steranu C30 v několika vzorcích z každého vrtného jádra v Kotlinském horizontu obsahuje nízké, ale detekovatelné množství biomarkeru demosponge steranu známého jako 24-isopropylcholestan (24-ipc)59,60 . Celkem byly sterany 24-ipc zaznamenány ve vzorcích hornin a ropy datovaných již do kryogénu (před >635 mil. let) v solné pánvi jižního Ománu a představují nejstarší lipidový biomarker dokládající výskyt metazoí59,60. Biomarkery 24-ipc v našich vzorcích měly buď přibližně o jeden řád nižší množství (vzhledem k celkovému poměru C27 a C30 steranů to bylo pouze 0,06-0,61 %; průměr = 0,22 %) ve srovnání s ediakarskými horninami a oleji z jižního Ománu (v průměru 1,7 %59), nebo byly u většiny vzorků pod detekčními limity kvůli zanedbatelnému množství.
Poměr izotopů dusíku a organického uhlíku
Izotopy dusíku mohou pomoci rozeznat relativní rovnováhu v cyklu dusíku a míru, do jaké byla fixace dusíku nebo neúplná denitrifikace významnou cestou ovlivňující bilanci živin dostupných pro mořská společenstva. Pokud diazotrofní bakterie fixují molekulární dusík v důsledku nedostatku fixovaného dusíku ve vodním sloupci, může to vést k hodnotám sedimentárních objemových izotopů dusíku blízkým 0 ‰61. To je v kontrastu s pozitivními izotopovými signaturami dusíku (v rozmezí +2 až +10 ‰) s modem +4 až +6 ‰ zjištěnými u neoproterozoických mořských sedimentů uložených v podmínkách, o nichž se předpokládá, že jsou plné dusičnanů, kde dusičnany prošly pouze částečnou denitrifikací62. Hodnoty izotopů dusíku pro všechny naše vzorky kromě jednoho odlehlého (tab. 1) pokrývají omezený kladný rozsah od +3,5 do +6,5 ‰, který se překrývá s modem pro pozdně neoproterozoické sedimentární horniny bohaté na organickou hmotu62. Naše data, omezená redoxními proxy ukazujícími, že ve vodním sloupci převládaly oxické podmínky, naznačují, že ve fondu rozpuštěného anorganického dusíku dominovaly dusičnany. Rozsah hodnot δ15N rovněž naznačuje, že fixace N2 nebyla primárním způsobem získávání dusíku pro primární producenty. Koloběhu dusíku spíše pravděpodobně dominovala recyklace ve vodním sloupci a organický N podléhal kvantitativní oxidaci na dusičnany, jak je tomu v moderním, proximálním mořském prostředí. Bentická denitrifikace a pohřbívání organického N by byly primárními pohlcovači rozpuštěného anorganického dusíku a rozsah hodnot δ15N naznačuje pouze omezenou roli neúplné denitrifikace ve vodním sloupci, která obvykle vede k významnému obohacení 15N63. Omezená variabilita hodnot δ15N proto pravděpodobně odráží δ15N dusičnanů advenovaných na platformu s malým vlivem omezené redukce dusičnanů ve vodním sloupci a fixace N2. Bez silného důkazu δ15N o fixaci dusíku předpokládáme, že dusík nebyl primární biolimitující živinou v epikontinentálních pánvích Baltiky. Izolace od říčních a eolických zdrojů fosforu a oxické podmínky v rozsáhlých mělkomorských epikontinentálních pánvích mohly zvýšit odstraňování autogenního fosforitu a stopových kovonosných fází, což vedlo k omezení živin, které omezovalo růst a produkci eukaryotických buněk.
Hodnoty izotopů celkového organického uhlíku (CTOC) se pohybují od -23,0 do -33,9 ‰, přičemž největší rozdíly byly pozorovány mezi jednotlivými lokalitami vrtných jader. Relativní obohacení 13C na izotopové poměry vyšší než cca -28 ‰ kontrastuje s údaji ze soudobých vrstev uložených v eutrofním prostředí otevřeného moře v Ománu30,31 . Námi uváděný rozsah je však obecně v souladu s rozsahem δ13CTOC pro jiné lokality z Baltiky uložené ve stejném časovém období11. Rozdíl mezi Baltikou a Ománem může poukazovat na skutečnost, že hodnoty δ13CTOC neodrážejí výhradně jednotnou sekulární změnu v koloběhu uhlíku v tomto časovém intervalu31,64. Rozsah δ13CTOC z Baltiky může spíše částečně odrážet mikrobiální ekologii s převahou bakterií, kterou naznačují jedinečné poměry lipidových biomarkerů. Malá velikost buněk, vysoký poměr plochy k objemu a pomalá rychlost růstu v oligotrofních podmínkách mohou zvýšit velikost frakcionace během autotrofie (εp)65,66, což vede k nízkým hodnotám δ13C.
Mezinformace pro více obohacené izotopové signatury 13C v rámci rozsahu uváděného11 pro pozdně ediakarskou sukcesi Baltiky je méně jasná. Možný příspěvek detritické, metamorfně pozměněné organické hmoty k prekambrickým sedimentárním sukcesím s nízkým obsahem TOC je možným mechanismem pro rozdíly mezi lokalitami v δ13CTOC64. Nicméně obsah TOC v našich vzorcích, přestože je nízký, je obecně vyšší než 0,10 hm. %, což neodhaluje žádné vztahy mezi obsahem TOC a hodnotami δ13C. Kromě toho naše vzorky obsahují tepelně nezralé organické látky a biomarkery lipidů, které neodpovídají převážně alochtonnímu zdroji uhlíku. Znaky δ13CTOC obohacené o více 13C mohou být výsledkem alternativních mechanismů asimilace uhlíku. Mechanismy koncentrace uhlíku nebo aktivní příjem bikarbonátu prokaryoty mohou vést k menším hodnotám εp a vyšším hodnotám δ13CTOC67,68. Vznik ediakarské bioty mohl významně rozšířit mořské potravní sítě a stimulovat nové cesty mikrobiální heterotrofie, včetně možného příspěvku komplexního koloběhu uhlíku v bentických mikrobiálních rohožích. S dodatečným přihlédnutím k potenciálně důležité roli rozpuštěné organické hmoty69 jako zdroje uhlíku odráží šíře hodnot δ13CTOC pravděpodobně řadu biogeochemických procesů koloběhu uhlíku, které mohou být jedinečné pro vyvíjející se ediakarské mořské prostředí a mohou souviset s jevem, pro který nemáme reprezentativní moderní analogy.
V raně triasových horninách z jižní Číny byly nalezeny starobylé sestavy lipidových biomarkerů a indikátory stabilních izotopů pro bentickou, mikrobiální produkci matů v tandemu s omezením fixního dusíku70. V důsledku masového vymírání na konci permu v Meishanu došlo k velkému nárůstu poměru hopanů a steranů (až na cca 60) doprovázenému silným posunem izotopové signatury Norg k hodnotám 0 až -2 ‰, což odpovídá bakteriální diazotrofii. To je spojeno s vysokým signálem 2-methylhopanů (2-methylhopanový index až 33 %) a výrazných methylalkanů, které ukazují na šíření bentických mikrobiálních rohoží. V rámci našeho souboru ediakarských dat nepozorujeme podobný trend ve vzorcích izotopů dusíku nebo biomarkerů, které by zjevně ukazovaly na významný vstup mikrobiálních rohoží, ačkoli v našich vzorcích byla zjištěna ojedinělá hodnota ochuzená o 15N (tab. 1). Silný bakteriální signál pozorovaný v našich baltických vzorcích tedy pravděpodobně není převážně signaturou bentických mikrobiálních rohoží.
Paleoenvironmentální výživa ediakarské bioty vs. biota. demosponges
Zjevné oligotrofní podmínky napříč epikrasovými a kontinentálními okrajovými pánvemi Baltiky, jak se od konce ediakaru do počátku kambria posouvala z vysokých do nízkých zeměpisných šířek, mohou souviset buď s omezenou advekcí relativně živinami bohatých hlubokých vod, nebo s vyčerpáním živin v důsledku asimilace a vymývání během transportu a usazování v těchto širokých, mělkomorských epikrasových pánvích (obr. 3). Tyto pánve byly epizodicky izolovány od oceánů a vyvinula se v nich hypersalinita (např. v době redkinské) a brakické podmínky (např. v době kotlinské71). Dlouhodobá tektonická stabilita měla za následek málo reliéfní topografii pozdně ediakarské Baltiky, velmi náchylnou k zaplavování a neefektivnímu přísunu fosforu pocházejícího ze zvětrávání. Na rozdíl od Baltiky odhalily biomarkerové studie vrstev ze superskupiny Huqf v Ománu eutrofní ekosystém, bohatý na mikrořasy30,31,59, ale bez ediakarské bioty, a to i v rozsáhlých výchozech v Ománských horách a v oblasti Huqf, v prostředí vnitřního až vnějšího šelfu3. Paleogeografie solné pánve v jižním Ománu byla rekonstruována pro pozdní neoproterozoikum ve vzdálenosti ~13° od rovníku na jižní polokouli, v podstatě podobně jako paleolituda Baltiky72.
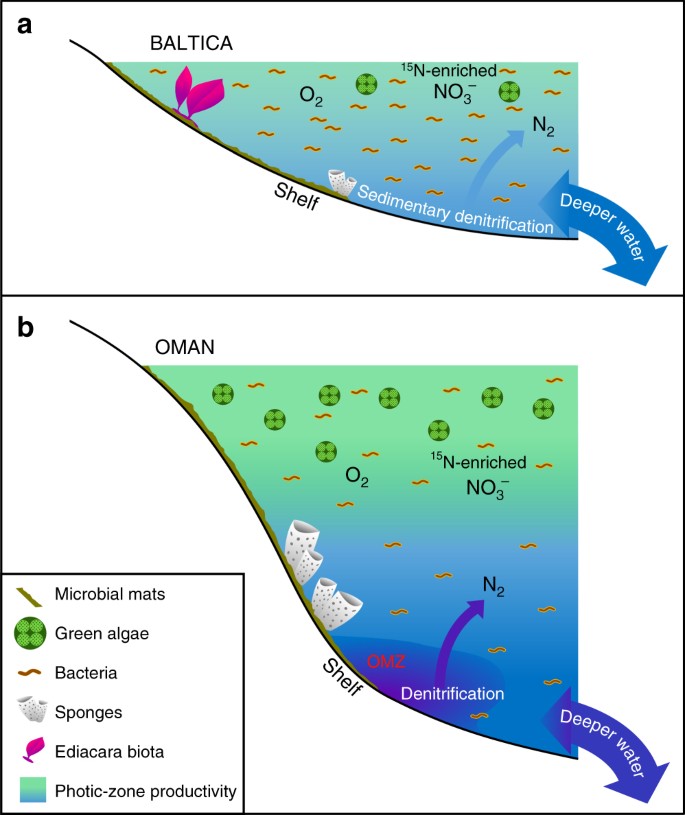
Významné rozdíly v málo produktivním vs. produktivním ediakarském mořském prostředí. Schémata jsou znázorněna pro rozsáhlé oligotrofní a mělkomorské okraje epikontinentálních pánví Baltiky, kde často převládala bakteriální produktivita a kde se dařilo ediakarské fauně měkkých těles a kde byly denitrifikace a anammox pravděpodobně omezeny na sedimenty; a b eutrofní a hlubší mořské šelfové prostředí solné pánve jižního Ománu, kde se dařilo zeleným řasám jako primárním producentům a hojně se vyskytovaly demospongie, ale fauna měkkých těl Ediacara nebyla výrazná (fosilie bioty Ediacara chyběly i v korelujících ediakarských výchozech v severním Ománu). Na produktivních kontinentálních okrajích pravděpodobně probíhala denitrifikace a anammox jak ve vodním sloupci, tak v sedimentech
Nejsou důkazy o trvalé a rozsáhlé anoxii na mělkých kontinentálních okrajích Baltiky a v jejich okolí během pozdního ediakaru11, a my se domníváme, že zde popsané oligotrofní podmínky byly způsobeny neefektivními terestrickými a hlubokovodními toky živin do těchto prostředí, které jsou v zásadě podobné podmínkám v oligotrofních prostředích moderních oceánských systémů. Geochemické zkoumání stopových prvků v Utkina Zavodu a přilehlých vrtných jádrech z oblasti Petrohradu skutečně naznačuje, že naše vzorky byly uloženy v oxických podmínkách73. Tuto interpretaci nezávisle podporují naše údaje z rozšířených distribucí hopanů (C31-C35), které s rostoucím počtem uhlíků prudce chvostovitě klesají v množství nad sloučeninami C31, což je charakteristické pro degradaci postranních řetězců bakteriohopanepolyolů během diageneze v oxických podmínkách29, a také nízkými vodíkovými indexy, měřenými pyrolýzou Rock-Eval (všechny nižší než 230 mg/g TOC a mnohé jsou nižší než 100 mg/g TOC) pro tyto nezralé vzorky v důsledku vzniku rekalcitrantního kerogenu oxidativní degradací a rekondenzací lipidově chudé primární biomasy v lokálně oxickém prostředí (tab. 1). Ediakarské mnohobuněčné organismy, které existovaly v těchto mělkomorských prostředích, musely mít k dispozici dostatek organických substrátů pro heterotrofii k uspokojení svých potravních potřeb spolu s dostatkem kyslíku a dalších živin k udržení svého metabolismu. Epikontinentální pánve Baltiky byly pravděpodobně trvaleji oxické než vysoce produktivní prostředí, která lemovala zóny kyslíkového minima, např. na středním až vnějším šelfu solné pánve Jižního Ománu, kde by respirace hojné planktonní biomasy udržovala nižší obsah rozpuštěného kyslíku pod fotickou zónou.
Ediakarská biota a další mnohobuněčné organismy žijící na pobřeží Baltiky by se musely vyrovnávat s měnícími se zdroji potravy (bakteriální vs. eukaryotické), protože tok živin se v čase měnil, včetně malých buněk a organického detritu v epikontinentálních pánvích, kde byly bakterie dominantními primárními producenty (obr. 3). Moderní společenstva korálových útesů přežívají v tropickém oligotrofním prostředí díky účinné recyklaci živin, včetně tvorby toku rozpuštěné organické hmoty (DOM), která pomáhá udržovat heterotrofii fauny v ekosystému útesu74. Vytvoření mořské trofické struktury s eukaryotickými mnohobuněčnými organismy, které se živí organickým detritem, muselo následovat po globální environmentální expanzi eukaryot do rozmanitých mořských prostředí, k níž došlo v průběhu toniánsko-kryogenního intervalu (cca 800-635 mil. let), jak bylo zjištěno ze záznamů biomarkerů15. Mohli bychom tedy očekávat, že v průběhu pozdního ediakaru rozeznáme důkazy o významných rozdílech ve struktuře mořských společenstev a trofické struktuře mezi jednotlivými lokalitami, přičemž místní bilance živin vybírala mikrobiální společenstva bohatá na eukaryota nebo chudá na eukaryota a postupná expanze mnohobuněčných organismů přidávala další rozměr složitosti na úrovni organismů a společenstev. Významný regionální kontrast v dostupnosti fosfátů a dalších živin v šelfovém prostředí je také očekávaným důsledkem heterogenní globální mořské redoxní struktury pro pozdní ediakar, před okysličením hlubokého oceánu38.
Dominance pikoplanktonní bakteriální produktivity a souvisejících produktů rozkladu rozpuštěné organické hmoty (DOM) v epikontinentálních pánvích Baltiky mohla udržovat mikrobiální smyčkový ekosystém souběžně s konvenční trofickou strukturou založenou na přibližně větších planktonních producentech50. To mohlo podporovat různé způsoby heterotrofie, včetně suspenzního krmení a případně osmotrofie, jako životaschopné potravní strategie pro některé rangeomorfy, houby a další pozdně ediakarské mnohobuněčné organismy18 ve spojení se vznikající aktivní (pohyblivou) heterotrofií19,20. Ve vodních systémech s omezeným obsahem živin zpravidla často dominuje malý jednobuněčný fytoplankton a heterotrofní plankton, přičemž v oligotrofních mořských prostředích současného oceánu49,51 převažují bakterie nad eukaryoty a udržují nízkou čistou biomasu. DOM je důležitým zdrojem organických živin a často řídí produktivitu a čistou biomasu v moderních oligotrofních tropických mořích, ale také kolimitaci dusíku a fosforu75. Podobně byl DOM pravděpodobně také důležitým substrátem pro udržení heterotrofních bakterií a mikrobiální smyčky v dávných oligotrofních prostředích. Součástí tohoto potravního řetězce s převahou bakterií mohly být bentické mikrobiální rohože, přičemž se předpokládá, že rohože přispěly k lepšímu uchování bioty Ediacara v mořském prostředí7. Zatímco u části našich vzorků byly zjištěny střední hodnoty 2-methylhopanového indexu (4-10 %, tab. 1), velmi nízké množství methylalkanů v poměru k n-alkanům (obr. 1) a pouze stopové a ojedinělé výskyty karotenoidů naznačují, že mikrobiální rohože nedominovaly primární produktivitě76 a že se dařilo planktonním bakteriím a odpovídajícím způsobem jejich produktům rozkladu DOM. Obecně nízké hodnoty vodíkového indexu (tab. 1; zejména u vzorků z Redkina), navzdory nízké teplotní zralosti vrstev a biomarkerových souborů, rovněž odpovídají organickému vstupu chudému na lipidy v převážně oxickém, mělkomorském okrajovém prostředí. Výskyt mikrobiálních matů měřený pouze ze sedimentologických textur nám v žádném případě nevypovídá o poměru eukaryot (mikrořas) a bakterií, ani nemůže omezit relativní podíl mikrobiálního planktonu na celkové primární produktivitě a výživě potravních sítí. Společenstva mikrobiálních rohoží, která často obsahují hojné množství eukaryot, a karbonáty skupiny Ara z jižního Ománu s výraznou trombolitickou a vrásčitou laminitovou facií jsou dobrým příkladem pozdně neoproterozoických prostředí s významným podílem mikrobiálních rohoží, která poskytují hojné steranové signály díky velkému podílu mikrořas30,59.
Velmi nízká úroveň biomarkerů 24-ipc naznačuje, že ačkoli byly demospongie někdy přítomny, byly v těchto prostředích řídké. To by mohlo naznačovat menší možnosti pro menší filtrující živočichy v konkurenci s biotou Ediacary v těchto nízkoprodukčních prostředích, nedostatečnou odolnost demospongí vůči energeticky náročnějším podmínkám mělkého moře nad základnou pouťových vln nebo lepší adaptabilitu hub na podmínky s nízkým obsahem kyslíku dynamicky udržované pod fotickou zónou v eutrofních prostředích (obr. 3). Celkově nebyl 24-ipc zjištěn ve většině vzorků redkinského horizontu, které mají také obecně nejvyšší poměry H/St z našeho souboru vzorků, což možná ukazuje na ekologickou změnu do doby uložení kotlinského horizontu. Alternativně je také pravděpodobné, že houby sice obývaly tato prostředí, ale neprodukovaly tyto diagnostické steroidní biomarkery v hojném množství, i když se to zdá méně pravděpodobné, protože steran 24-ipc patří mezi nejčastěji detekované sloučeniny steranu C30 v ediakarských vrstvách a olejích30,3133,59,60 a je detekovatelný i v podskupině našich vzorků (tab. 1).
Přestože trvalá oligotrofní mořská prostředí naznačená našimi údaji představují lokální podmínky v ediakarských oceánech, pravděpodobně nebyla neobvyklá pro prekambrické mělkomorské mořské cesty (obr. 3). Převážná většina předchozích ediakarských studií biomarkerů byla provedena na organicky bohatých sedimentárních horninách usazených v eutrofním prostředí a jejich ropných produktech, které obecně poskytují biomarkerové soubory odpovídající významnému podílu mikrořas44. Naše výsledky zdůrazňují význam studia širší škály depozičních prostředí, včetně vrstev chudých na organickou hmotu s vhodnou termální zralostí a různou litologií15,3034,59, aby bylo možné získat přesnější představu o rozsahu heterogenity mořské chemie a ekologie v jednotlivých lokalitách. Navzdory postupujícímu provzdušňování oceánu a zvýšenému chemickému zvětrávání a přísunu živin během rozpadu Rodinie a v průběhu ediakarského období77,78 udržovala oceánská heterogenita různé mořské chemické podmínky, včetně živinami chudého, ale obyvatelného prostředí, které podporovalo adaptaci, konkurenci a evoluci metazoí v systému globálního oceánu (obr. 3). Zatímco eutrofní mořské šelfy hostily demospongy, ale často postrádaly ediakarskou biotu, přičemž výrazným příkladem je solná pánev v jižním Ománu (a s ní korelující silné ediakarské výchozy severněji v Ománu); naopak mělké, oxické a méně produktivní epikontinentální mořské cesty byly kolonizovány ediakarskou biotou přednostně před demospongy, přestože DOM a další organický detrit byly lokálně dostupné pro výživu. Zda metabolické požadavky nebo selekční tlak prostředí omezily ediakarskou měkkotělesnou mnohobuněčnou biotu na tato prostředí, zůstává nejisté; naše studie však zdůrazňuje, že ediakarské oligotrofní prostředí hrálo potenciálně klíčovou roli v evoluci makroskopických mnohobuněčných organismů a ekologii mořských společenstev.