Szokatlanul alacsony termikus érettség
A termikus érettség az egyetlen legfontosabb tényező, amely befolyásolja az ősi üledékes biomarkerek megőrzését, mivel a szerves molekulák a fokozatos betemetés során és a növekvő termikus stressz hatására szerkezeti és sztereokémiai változásokra hajlamosak29. A megfelelő termikus érettségű prekambriumi kőzetek (olajablak-érettség vagy annál alacsonyabb) előfeltételei a robusztus és elsődleges biomarker lipidegyüttesek megőrzésének. A többszörös hopán és szterán biomarker sztereoizomer arányok, valamint a Rock-Eval pirolízisből származó alacsony Tmax értékek (többnyire a 417-433 °C tartományban, átlagosan 426 °C) független bizonyítékai azt mutatják, hogy a jelen tanulmányban szereplő kőzetek messze a legmodernebb szerves geokémiai módszerekkel eddig elemzett, termikusan legéretlenebb ediakari kőzetek (1. táblázat). Bár kőzeteink üledékes diagenezisen mentek keresztül, nem léptek át jelentősen az olajablakba, eltekintve a moldovai Podillya-medencéből származó, kissé érettebb 16PL feltárási mintáktól, amelyek még mindig alkalmasak az elemzésre (az olajablak korai-közepes érettsége és szerves szennyeződések nyilvánvaló jelei nélkül). A készlet nagy része ekkor valószínűleg nem volt kitéve 50 °C-ot meghaladó temetkezési hőmérsékletnek, így a mintáink nagyon éretlenek. Az alacsony termikus érettséget és szinogénséget alátámasztó további molekuláris bizonyítékok közé tartozik (i) a policiklusos biomarker alkánok dominanciája az n-alkánokkal szemben a kőzetkivonatokban (2. ábra), (ii) a stabil 17β,21β(H)- és 17α,21β(H)-stereokémiai konfigurációval rendelkező, nagyobb mennyiségben előforduló hopánoktól elkülöníthető 17β,21α(H)- és 17α,21β(H)-stereokémiai konfigurációval rendelkező, kimutatható mennyiségű hopánok túlélése (1. táblázat, Kiegészítő ábrák. 1. és 2. ábra), (iii) a C22 és C27 közötti n-alkánok között a páros és a páratlan közötti páratlan preferencia (2. ábra), mivel az n-alkánok csak a katagenezis előtt mutatnak szénszámpreferenciát38 , és (iv) termikusan éretlen hopán és szterán biomarkerek előállítása a (nem oldódó) kerogén fázisból katalitikus hidropilízis segítségével (2. kiegészítő ábra). Az üledékes szerves anyag alacsony termikus érettsége a mintáinkban összhangban van az üledékes alteráció korábban publikált mutatóival, beleértve a konodont és akritarcha alterációs indexeket, valamint a Baltica ediakari és paleozoikus üledékes kőzetek korábbi Rock-Eval pirolízis és biomarker vizsgálataival36,39,40 , valamint az agyagmineralológiával41. Kritikus, hogy olyan környezetben, ahol a termikus érettség ilyen alacsony, az elsődleges δ15N és δ13C jelek jelentős változásának mechanizmusai nagyrészt hiányoznak42.

A kivonható alifás szénhidrogének eloszlása egy reprezentatív minta esetében. a A kivonható alifás szénhidrogének teljes ionkromatogramja (TIC) a Redkino Horizonból származó Lugovoe #13-73 m számú Lugovoe #13-73 m esetében. Az n-alkán sorozat, a prisztán (Pr), a fitán (Ph) és a C27-C34 hopének (az összes szénatomszámukkal és a C-17, C-21 és C-22 sztereokémiával jelölve, pl. C31αβR) jelölve vannak; ** a C29 és C30 hopéneket jelöli. Figyeljük meg a hopánok bőségének dominanciáját a többi alkánvegyület-osztályhoz képest. b A részleges 85-Da ionkromatogram az n-alkán-sorozat dominanciáját mutatja, amely a C22-C27 tartományban a páros és a páratlan szénatomszámok közötti egyértelmű preferenciát mutat, a metilalkánokkal szemben
Lipid biomarkerek összetételének mintázatai
A fő (C27-C35) hopánok és a fő (C27-C29) szteránok gyakorisági arányát gyakran használják a baktériumok és eukarióta forrásszervezetek ősi vízi ökoszisztémába történő bevitelének egyensúlyának értékelésére. A hopánok hopanoidokból származó molekuláris fosszíliák, amelyek a legkülönbözőbb baktériumcsoportok által szintetizált sejtmembrán-lipidek. Hasonlóképpen, a szteránok szterol prekurzorokból származnak, amelyeket szinte kizárólag eukarióták állítanak elő43. A mintáink hopán/sterán (H/St) arányai feltűnően nagy értéktartományt fednek le, 1,6-tól 119,2-ig (1. táblázat). Összehasonlításképpen, a szerves anyagokban gazdag neoproterozoikus kőzetekből és olajokból származó H/St arányok jellemzően 0,5 és 2,030,31,32,33 közötti szűk tartományba esnek. Bár egyértelmű különbség van a fiatalabb Kotlin-horizont (átlagos H/St 8,9) és az idősebb Redkino-horizont (átlagos H/St 42,9) értékei között, a legtöbb minta értékei szokatlanul magasak, és a baktériumok rendellenesen magas hozzájárulására utalnak.
A késő neoproterozoikumra az eukarióta algák ökológiailag jelentős összetevői és fő termelői voltak számos tengeri ökoszisztémának15,34,44. Az eltérés az eukarióta algák globálisan jelentős hozzájárulása között a késő neoproterozoikumban, és mégis a szterán biomarkerek alacsony szintje között ezekben a mintákban azt jelzi, hogy az eukarióta bőséget valamilyen helyi meghatározó tényezőnek kell lennie. A legegyszerűbb magyarázat a mintáinkban talált rendkívül magas hopán/szterán arányokra az alacsony teljes szerves széntartalom (TOC) és az alacsony hidrogénindex (HI) mellett (1. táblázat) az, hogy ezek a rétegek oligotróf (azaz erősen tápanyag-korlátozott) környezetben rakódtak le, ahol a baktériumok kiszorították az algákat. Bár nehéz modern analógiákat találni a magas tengerszint-állás során kialakult ősi epikratonikus tengerekre, korábban már találtak párhuzamos megfigyeléseket a magasabb (akár egy nagyságrenddel) hopán/szterán arányokról az ordovícium-szilur epikontinentális tengerekben lerakódott, szerves anyagokban szegény és szerves anyagokban gazdag üledékes kőzetek esetében45.
Az alacsony TOC-tartalmat tipikusan a szerves anyag korlátozott lerakódása okozhatja alacsony termelékenységű környezetben, a szerves anyag alacsony megőrzési potenciálja a vízoszlopban vagy az üledékekben, vagy a magas sziliklasztikus fluxussal történő hígulás. A modern óceánokban a termelékenységet leggyakrabban az alapvető tápanyagok – nitrogén, foszfor, szilícium és vas – alacsony szintje korlátozza46. A nitrogén, a foszfor és a vas korlátozott a nyílt óceáni régiókban, ahol a feláramlás, a porbevitel és a part menti lefolyás nem szállít elegendő mennyiségű tápanyagot. A modern oligotróf környezetben a baktériumok és az eukarióta biomassza aránya magasabb, mint az eutróf vagy mezotróf környezetben. Érdekes módon a moldvai Redkino horizontból származó Podillya 16PL feltárási minták foszforittartalmú iszapkövek, és lényegesen alacsonyabb hopán/szterán arányt (8,1:11,5) adnak, mint az 1. táblázatban szereplő többi Redkino minta (bár még mindig jelentősen magasabb, mint a Dél-Omani Sómedencében vagy a szerves anyagokban gazdag fanerozoikus üledékekben találtak, amelyek jellemzően egy szűk, 0.5-2,0 közötti tartományba esnek), ami valószínűleg arra utal, hogy a megnövekedett foszfát elérhetőség kedvezően befolyásolhatta a helyben talált eukarióta-baktérium arányt (tekintve, hogy a nitrogénizotópos jelek nagyrészt változatlanok, lásd a következő szakaszt). A foszfor (P) tartalom, valamint a P/Fetotal és P/Al arányok egyébként általában alacsonyak a Podillya, Ukrajna és Észtország47 késő ediakari üledékeiben; kivéve ezt a foszforitgumókkal jelölt rétegtani szintet. Hasonlóan alacsony (~0,01 és ~0,1 tömegszázalék közötti) P-tartalomról számoltak be a Kotlin és Redkino sziliklasztos kőzetekben a Kelet-európai Platform északkeleti pereméről származó fúrómagból11.
A Baltikát átszelő epikontinentális tengerek széles és sekély topográfiája fenntarthatta a foszfor- vagy más tápanyagkorlátozásokat a peremvidéki környezetben, ami a foszfátnak az oxikus felszíni vizekben a vasásványokkal való autogén kicsapódásának és számos elemnek a part menti üledékekben való megkötésének köszönhető48. Ha a foszfát volt a korlátozó tápanyag, akkor a biológiailag hozzáférhető foszfor növelése fokozta volna a helyi elsődleges termelést, és kedvezőbb növekedési feltételeket biztosított volna a nagyobb méretű egysejtű fitoplankton számára49,50,51 . Ami a modern óceáni rendszert illeti, a tengeri pikocianobaktériumok, a Prochlorococcus és a Synechococcus elismerten uralják a fitoplankton sejtszámát és biomasszáját az oligotróf trópusi és szubtrópusi óceáni környezetben, beleértve az óceáni felszíni vizek foszfátlimitált oligotróf régióit52. A Prochlorococcus és a heterotróf SAR11 (Pelagibacter) számos adaptációnak köszönhetően virágzik, beleértve a kis genom és az alacsony replikációs sebesség révén alacsony energiaköltségeket, a kisebb sejtméretek révén nagyobb felület-térfogat arányt, valamint a tápanyag-felhasználás maximalizálása érdekében további sejtfelvételi funkciókat53. Számos baktérium képes a tápanyaghiányos tápanyagokat is helyettesíteni, pl. a foszforban szegény környezetben a foszfortartalmú lipidek helyett a szulfolipidek felhasználásával54 , vagy alternatív szubsztrátok felhasználásával, pl. nitrogénmegkötés útján a légköri N2-ből származó nitrogén felhasználásával55 , a tápanyagstressz enyhítése érdekében. A tengeri bakterioplankton oligotróf környezetben való túlélésére vonatkozó, nemrégiben készült tanulmány56 , ahol a rendelkezésre álló foszfát korlátozott, arra utal, hogy a polifoszfát-anyagcsere fontos szerepet játszik a tengeri oligotrófoknál. Az oligotróf környezetben versenyző kiválasztott eukarióták jellemzően kisméretű pikoeukarióták, amelyek táplálkozási igényeiket mixotrófia57 révén egészíthetik ki. A pikoeukarióták és a cianobaktériumok biomassza aránya fokozott tápanyagellátás esetén általában növekszik49. A foszfor és más biolimitáló tápanyagok lehetséges hatása a primer termelékenység és a tengeri közösségek szerkezetének mérséklésében az ediakari epeirikus tengerekben további vizsgálatokat igényel.
A C29-steránok gyakorisága kivétel nélkül nagyobb, mint a megfelelő C27- vagy C28-steránoké valamennyi helyszínünkön (1. táblázat). A C29-steránok túlsúlya a C27- és C28-steránokkal szemben valószínűleg a zöld algák dominanciáját jelzi az eukarióta fitoplanktonközösségen belül44,58 . Ez a jellemző a legtöbb korábbi ediakaráni biomarker-vizsgálat során is megfigyelhető volt15,30,31,32,33,34,35,36,44 . Figyelemre méltó, hogy a Kotlin Horizon egyes fúrómagjaiból származó több mintában a C30-sterán eloszlása alacsony, de kimutatható mennyiségben tartalmazza a 24-izopropil-kolesztán (24-ipc)59,60 néven ismert demoszpóniás sterán biomarkert. Összesen 24-ipc-steránokról számoltak be kőzet- és olajmintákban, amelyek egészen a kriogén korig (>635 Ma ezelőtt) nyúlnak vissza a Dél-Omani Sómedencében, és ezek jelentik a metazoák legrégebbi lipid biomarkereit59,60 . A 24-ipc biomarkerek a mintáinkban vagy körülbelül egy nagyságrenddel kisebb mennyiségben fordultak elő (a teljes C27-C30 szterán arányhoz viszonyítva ezek aránya csak 0,06-0,61% volt; átlag = 0,22%), mint a Dél-Ománból származó ediakarikus kőzetekben és olajokban (átlagosan 1,7%59 ), vagy a minták többségében elhanyagolható mennyiségük miatt a kimutatási határérték alatt voltak.
A nitrogén és szerves szén izotóparányok
A nitrogénizotópok segíthetnek megkülönböztetni a nitrogénciklus relatív egyensúlyát, és azt, hogy a nitrogénkötés vagy a nem teljes denitrifikáció milyen mértékben volt jelentős útvonal a tengeri közösségek számára elérhető tápanyagegyensúly befolyásolásában. Ha a diazotróf baktériumok molekuláris nitrogént kötnek meg, mivel a vízoszlopban nincs kötött nitrogén, ez 0‰61 közeli üledéki nitrogénizotóp-értékeket eredményezhet. Ez ellentétben áll a pozitív nitrogénizotóp-jelzésekkel (+2 és +10‰ közötti tartományban), amelyek +4 és +6‰ közötti értéket mutatnak a neoproterozoikus tengeri üledékekben, amelyek a feltételezések szerint nitrátban gazdag körülmények között rakódtak le, ahol a nitrát csak részleges denitrifikáción ment keresztül62. A nitrogénizotóp-értékek a mi mintáinkban egy kivételével (1. táblázat) egy korlátozott pozitív tartományt fednek le, +3,5 és +6,5‰ között, ami átfedésben van a késő neoproterozoikum szerves anyagban gazdag üledékes kőzetekre vonatkozó módusszal62. A redoxi mutatók alapján, amelyek arra utalnak, hogy a vízoszlopban oxikus körülmények uralkodtak, adataink arra utalnak, hogy a nitrát dominált az oldott szervetlen nitrogénkészletben. A δ15N értékek tartománya arra is utal, hogy az elsődleges termelők számára nem a N2-fixálás volt a nitrogénszerzés elsődleges módja. A nitrogénciklusban valószínűleg inkább a vízoszlopban történő újrahasznosítás dominált, és a szerves N mennyiségi oxidációnak volt kitéve nitráttá, ahogyan az a modern, proximalis tengeri környezetben is történik. A benti denitrifikáció és a szerves N eltemetése lehetett az oldott szervetlen nitrogén elsődleges nyelője, és a δ15N értékek tartománya arra utal, hogy a nem teljes vízoszlopbeli denitrifikációnak csak korlátozott szerepe volt, ami általában jelentős 15N-dúsulást eredményez63. A δ15N-értékek korlátozott változékonysága ezért valószínűleg a platformra szállított nitrát δ15N-jét tükrözi, a vízoszlop korlátozott nitrátredukciójának és N2-fixálásának csekély befolyása mellett. A nitrogénmegkötés erős δ15N bizonyítékainak hiányában feltételezzük, hogy a nitrogén nem volt az elsődleges biolimitáló tápanyag a Baltica epikontinentális medencéiben. A folyami és eolikus foszforforrásoktól való elszigetelődés, valamint a széles, sekélytengeri epikontinentális medencék oxikus viszonyai fokozhatták az autogén foszforit és a nyomfémtartalmú fázisok eltávolítását, ami az eukarióta sejtek növekedését és termelését korlátozó tápanyaglimitációt eredményezett.
A teljes szerves szén (CTOC) izotópértékei -23,0 és -33,9‰ között változnak, a legnagyobb különbségek a különböző fúrási pontok között figyelhetők meg. A kb. -28‰-nél magasabb izotóparányokhoz viszonyított relatív 13C-dúsulás ellentétben áll az ománi eutróf, nyílt tengeri környezetben lerakódott kortárs rétegekből származó adatokkal30,31 . Az általunk közölt tartomány azonban általában összhangban van az ugyanabban az időszakban lerakódott δ13CTOC tartományával más balticai lelőhelyeken11. A Baltica és Omán közötti különbség rávilágíthat arra, hogy a δ13CTOC-értékek nem kizárólag a szénciklus egységes, szekuláris változását tükrözik ebben az időintervallumban31,64 . A balticai δ13CTOC-tartomány inkább a baktériumok által dominált mikrobiális ökológiát tükrözheti, amire az egyedi lipid biomarker-arányok utalnak. A kis sejtméret, a nagy felület-térfogat arány és a lassú növekedési sebesség oligotróf körülmények között növelheti a frakcionálás nagyságát az autotrófia során (εp)65,66 , ami alacsony δ13C értékeket eredményez.
A késő ediakaráni Baltica szukcesszióról jelentett11 tartományon belül a 13C-vel dúsabb izotópos jelek mechanizmusa kevésbé világos. A detritális, metamorfikusan átalakult szerves anyag lehetséges hozzájárulása a prekambriumi, alacsony TOC-tartalmú üledékes szukcessziókhoz egy lehetséges mechanizmus a δ13CTOC64 helyszínek közötti eltérésekhez. A mintáink TOC-tartalma azonban, bár alacsony, általában nagyobb, mint 0,10 tömegszázalék, ami nem mutat összefüggést a TOC-tartalom és a δ13C-értékek között. Továbbá, a mintáink termikusan éretlen szerves anyagot és biomarker lipidmintázatot tartalmaznak, ami nincs összhangban a főként allochton szénforrással. A 13C-vel dúsabb δ13CTOC jelek a szén asszimiláció alternatív mechanizmusainak eredményei lehetnek. A szénkoncentrációs mechanizmusok vagy a prokarióták aktív bikarbonátfelvétele kisebb εp-értékeket és magasabb δ13CTOC-értékeket eredményezhet67,68 . Az ediakarán biota megjelenése jelentősen kibővíthette a tengeri táplálékhálózatokat, és a mikrobiális heterotrófia új útjait serkentette, beleértve a komplex szénciklusok lehetséges hozzájárulását a benti mikrobiális szőnyegekben. Az oldott szerves anyag69 mint szénforrás potenciálisan fontos szerepének további figyelembevételével a δ13CTOC-értékek széles skálája valószínűleg olyan biogeokémiai szénciklus-folyamatokat tükröz, amelyek egyedülállóak lehetnek a fejlődő ediakari tengeri környezetben, és olyan jelenséghez kapcsolódhatnak, amelyre nincsenek reprezentatív modern analógjaink.
A dél-kínai kora-triász kőzetekben70 találtak ókori lipid biomarker-összetételeket és a fix nitrogénlimitációval párhuzamos benti, mikrobiális szőnyegtermelés stabil izotópos indikátorait. A perm végi tömeges kihalást követően Meishanban a hopán/szterán arányok nagymértékű megugrása (kb. 60-ig) a Norg izotóp szignatúra erős eltolódásával együtt 0 és -2‰ közötti értékekre változott, ami összhangban van a bakteriális diazotrófiával. Ehhez társul a 2-metilhopánok (2-metilhopán-index akár 33%) és a jellegzetes metilalkánok magas jele, ami a benti mikrobiális matracok elszaporodására utal. Nem figyelhető meg hasonló tendencia a nitrogénizotópos szignatúrában vagy a biomarkerek mintázatában, ami nyilvánvalóan jelentős mikrobiális szőnyegbevitelre utalna az ediakari adatsorunkon belül, bár a mintáinkban találtunk egy egyedi 15N-hiányos értéket (1. táblázat). Ezért a Baltica-mintáinkban megfigyelt erős bakteriális jel valószínűleg nem túlnyomórészt a benti mikrobiális szőnyegek jele.
Paleoenvironmental sustenance of Ediacara biota vs. Demosponges
A Baltica epikratonikus és kontinentális peremmedencéiben uralkodó nyilvánvalóan oligotróf körülmények, ahogy a Baltica a magas szélességektől az alacsony szélességek felé sodródott a késő ediakarumtól a korai kambriumig, vagy a viszonylag tápanyagban gazdag mélyvizek korlátozott advekciójával, vagy a tápanyagfogyással hozhatók összefüggésbe, amely az asszimiláció és a dögeltakarítás eredménye a szállítás és lerakódás során e széles, sekélytengeri epikontinentális medencékben (3. ábra). Ezek a medencék epizodikusan elszigetelődtek az óceánoktól, és hipersós (pl. a Redkino-időszakban) és brakkos (pl. a Kotlin-időszakban71) állapotokat alakítottak ki. A hosszú távú tektonikai stabilitás a késő ediakaráni Baltica alacsony domborzatú domborzatát eredményezte, amely rendkívül érzékeny volt az elöntésekre és az időjárásból származó foszfor nem hatékony ellátására. Balticával ellentétben az ománi Huqf szupercsoportból származó rétegek biomarker-vizsgálatai eutrofikus, mikroalgákban gazdag ökoszisztémát mutattak ki30,31,59 , de hiányzott az Ediacara élővilága, még az Omán-hegység és a Huqf régió kiterjedt feltárásaiban is, a belső és külső polcok közötti környezetben3. A Dél-Omán Sómedence paleogeográfiáját a késő neoproterozoikumra vonatkozóan a déli féltekén az Egyenlítőtől ~13°-ra rekonstruálták, ami nagyjából a Baltica72 paleolátushoz hasonló.
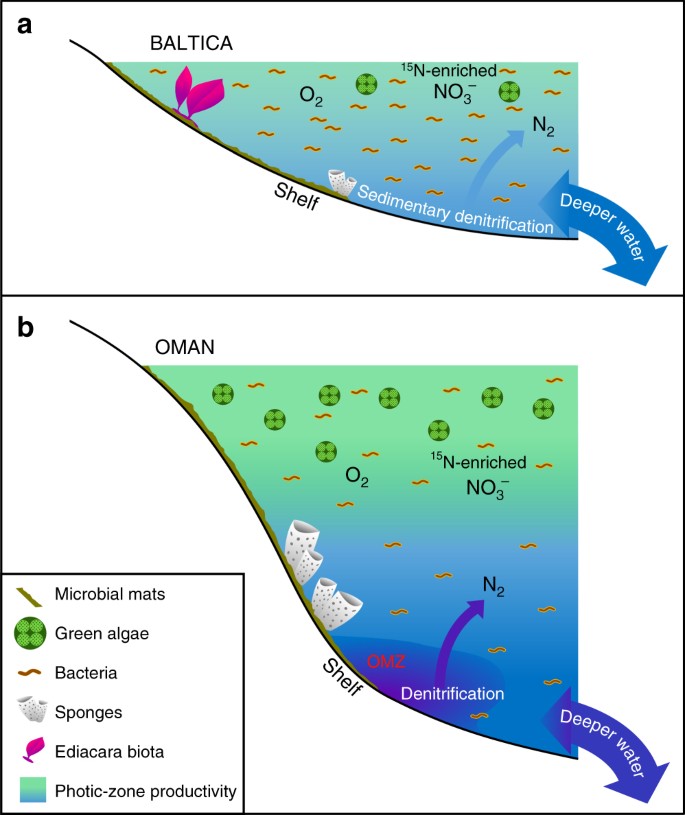
Nagy különbségek az alacsony termelékenységű vs. termelékeny ediakari tengeri környezetekben. Sematikus ábrák láthatók egy kiterjedt oligotróf és sekélytengeri epikontinentális medence peremére a Baltikumban, ahol gyakran a bakteriális termelékenység dominált, ahol az Ediacara lágytestű fauna virágzott, és ahol a denitrifikáció és az anammox valószínűleg az üledékekre korlátozódott; és b a Dél-Omán sós medencéjének eutróf és mélyebb tengeri talapzatai, ahol a zöld algák elsődleges termelőként virágoztak, és a demospongák bőségesek voltak, de az Ediacara lágy testű fauna nem volt kiemelkedő (az Ediacara biota fosszíliái szintén hiányoznak az Észak-Ománban található korrelatív ediakarikus feltárásokból). A produktív kontinentális peremeken a denitrifikáció és az anammox valószínűleg mind a vízoszlopban, mind az üledékekben előfordult
Nincs bizonyíték arra, hogy a Baltika sekély kontinentális peremein és azok környékén a késő ediakarán idején tartós és kiterjedt anoxia állt volna fenn11, és azt javasoljuk, hogy az itt leírt oligotróf körülményeket az ide irányuló nem hatékony szárazföldi és mélyvízi tápanyagáramlás okozta, amely nagyjából hasonló a modern óceáni rendszerek oligotróf környezetében tapasztaltakhoz. Az Utkina Zavod és a Szentpétervár környéki szomszédos fúrómagok nyomelem-geokémiai vizsgálata valóban arra utal, hogy a mintáink oxikus körülmények között rakódtak le73. Ezt az értelmezést egymástól függetlenül alátámasztják a kiterjesztett hopán (C31-C35) eloszlásokra vonatkozó adataink, amelyek a C31 vegyületek felett a szénszám növekedésével meredeken csökken a gyakoriság, ami a bakteriohopánpoliolok oldallánc-bomlására jellemző az oxikus körülmények közötti diagenezis során29, valamint a Rock-Eval pirolízissel mért alacsony hidrogénindexek (mindegyik 230 mg/g TOC alatt van, és sokan 100 mg/g TOC alatt vannak) ezekre az éretlen mintákra vonatkozóan, ami a helyileg oxikus környezetben a lipidszegény primer biomassza oxidatív lebomlásán és rekondenzációján keresztül történő rekalcitráns kerogén kialakulásának köszönhető (1. táblázat). Az e sekélytengeri környezetben létező ediakari többsejtű élőlényeknek elegendő szerves szubsztrátummal kellett rendelkezniük a heterotrófiához, hogy kielégítsék táplálkozási szükségleteiket, valamint elegendő oxigénnel és más tápanyagokkal az anyagcseréjük fenntartásához. A Baltica epikontinentális medencéi valószínűleg tartósan oxikusabbak voltak, mint az oxigénminimum-zónákat szegélyező, rendkívül termékeny környezetek, mint például a Dél-Omániai Sómedence középső és külső peremén, ahol a bőséges planktonikus biomassza légzése a fotikus zóna alatt alacsonyabb oldott oxigénszintet tarthatott fenn.
A Baltica partjainál élő Ediacara-biotának és más többsejtű szervezeteknek változó (bakteriális vs. eukarióta) táplálékforrásokkal kellett megbirkózniuk, ahogy a tápanyagáramlás az idők folyamán változott, beleértve a kis sejteket és a szerves detrituszt az epikontinentális medencékben, ahol a baktériumok voltak a domináns elsődleges termelők (3. ábra). A modern korallzátony-közösségek a trópusi oligotróf környezetben a tápanyagok hatékony újrahasznosításának köszönhetően maradnak fenn, beleértve az oldott szervesanyag-áramlást (DOM), amely segít fenntartani a fauna heterotrófiáját a zátony ökoszisztémán belül74. A szerves törmelékkel táplálkozó eukarióta többsejtű szervezeteket tartalmazó tengeri trofikus struktúra kialakulásának az eukarióták globális szintű környezeti terjeszkedése után kellett bekövetkeznie, amely a tonik-kriogén intervallumban (kb. 800-635 Ma) zajlott, amint azt a biomarker-feljegyzésekből megállapíthatjuk15. Ezután arra számíthatunk, hogy a tengeri közösségek és a trofikus struktúra jelentős helyenkénti különbségeinek bizonyítékait fedezhetjük fel a késő ediakarán időszak során, ahol a helyi tápanyagegyensúly eukariótákban gazdag vagy eukariótákban szegény mikrobiális közösségeket választott, és ahol a többsejtűek fokozatos elterjedése a komplexitás egy újabb dimenzióját adta hozzá a szervezetek és a közösségek szintjén. A foszfát és más tápanyagok elérhetőségének jelentős regionális kontrasztja a talapzat környezetében szintén a heterogén globális tengeri redoxszerkezet várható következménye a késő ediakaránban, a mély óceán oxigéndúsulása előtt38.
A pikoplankton baktériumtermékenység és a kapcsolódó oldott szervesanyag (DOM) lebontási termékek dominanciája a Baltica epikontinentális medencéiben a hagyományos, nagyobb méretű planktonikus termelők köré épülő trofikus struktúrával párhuzamosan fenntarthatott egy mikrobiális hurok ökoszisztémát50. Ez kedvezhetett a heterotrófia különböző módjainak, beleértve a szuszpenziós táplálkozást és esetleg az ozmotrófiát, mint életképes táplálkozási stratégiát egyes rangeomorfok, szivacsok és más késő ediakari többsejtű szervezetek18 számára, a kialakuló aktív (mozgó) heterotrófiával együtt19,20 . A tápanyag-korlátozott vízi rendszerekben általában a kis egysejtű fitoplankton és a heterotróf plankton dominál, a baktériumok túlszárnyalják az eukariótákat, és a modern óceánok oligotróf tengeri környezetében alacsony nettó biomasszát tartanak fenn49,51 . A DOM a szerves tápanyagok fontos forrása, és gyakran szabályozza a termelékenységet és a nettó biomasszát a modern oligotróf trópusi tengerekben, de a nitrogén- és foszfor-ko-limitáció is ilyen75. Hasonlóképpen, a DOM valószínűleg fontos szubsztrát volt a heterotróf baktériumok és egy mikrobiális kör fenntartásához az ősi oligotróf környezetben. A benti mikrobiális szőnyegek ennek a baktériumok által dominált táplálékhálózatnak az egyik összetevői lehettek, és a szőnyegalapok hozzájárulhattak az Ediacara élővilágának tengeri környezetben való megőrzéséhez7. Míg a 2-metilhopán-index mérsékelt értékeit találtuk a mintáink egy részhalmazában (4-10%, 1. táblázat), a metilalkánok nagyon alacsony előfordulása az n-alkánokhoz képest (1. ábra), és a karotinoidok csak nyomokban és szórványosan fordulnak elő, arra utal, hogy a mikrobiális matracok nem domináltak az elsődleges termelékenységben76 , és hogy a planktonikus baktériumok és ennek megfelelően a DOM-bontási termékeik virágoztak. Az általában alacsony hidrogénindex-értékek (1. táblázat; különösen a Redkino minták esetében), a rétegek és a biomarker-összetételek alacsony termikus érettsége ellenére, szintén összhangban vannak a lipidszegény szervesanyag-bevitellel a főként oxikus, sekélytengeri marginális környezetben. A mikrobiális szőnyegek előfordulása, amelyet csak a szedimentológiai textúrák alapján mérünk, semmiképpen sem mond semmit az eukarióták (mikroalgák) és a baktériumok közötti egyensúlyról, és a mikrobiális planktonnak az általános primer termelékenységhez és a táplálékhálózat fenntartásához való relatív hozzájárulását sem tudják korlátozni. A gyakran bőséges eukariótákat tartalmazó mikrobiális szőnyegközösségek és a Dél-Ománból származó Ara-csoport karbonátjai, amelyek kiemelkedő trombolitikus és ráncos laminit fáciesekkel rendelkeznek, jó példa a jelentős mikrobiális szőnyeg-hozzájárulással rendelkező késő neoproterozoikus környezetekre, amelyek a mikroalgák nagy hozzájárulása miatt bőséges szteránjeleket adnak30,59.
A 24-ipc biomarkerek rendkívül alacsony szintje arra utal, hogy bár a demospongák néha jelen voltak, ezekben a környezetekben ritkák voltak. Ez azt jelezheti, hogy ezekben az alacsony termelékenységű környezetekben kevesebb lehetőségük van az Ediacara élővilágával versengő kisebb szűrőtáplálkozó állatoknak, a demospongák elégtelen ellenálló képessége az energikusabb sekélytengeri körülményekkel szemben a szépvízi hullámbázis felett, vagy a szivacsok jobb alkalmazkodóképessége az eutróf környezetben a fotikus zóna alatt dinamikusan fenntartott alacsony oxigénszintű körülményekhez (3. ábra). Összességében a 24-ipc nem volt kimutatható a Redkino-horizont legtöbb mintájában, amelyek általában a legmagasabb H/St-aránnyal is rendelkeznek a mintasorozatunkban, ami talán ökológiai változást jelez a Kotlin-horizont lerakódási idejére. Másik lehetőségként az is elképzelhető, hogy a szivacsok ugyan benépesítették ezeket a környezeteket, de nem termelték nagy mennyiségben ezeket a diagnosztikus szteroid biomarkereket, bár ez kevésbé tűnik valószínűnek, mivel a 24-ipc-sterán az ediakaráni rétegekben és olajokban30,3133,59,60 leggyakrabban kimutatott C30-sterán vegyületek közé tartozik, és a mi mintáink egy részhalmazában is kimutatható (1. táblázat).
Míg az adataink által sugallt tartós oligotróf tengeri környezetek az ediakari óceánok lokális viszonyait képviselik, valószínűleg nem voltak szokatlanok a prekambriumi sekélytengeri tengerszorosokban (3. ábra). A korábbi ediakari biomarker-vizsgálatok túlnyomó többségét eutróf környezetben lerakódott, szerves anyagokban gazdag üledékes kőzeteken és azok kőolajtermékein végezték, amelyek általában olyan biomarker-összetételeket eredményeznek, amelyek jelentős mikroalga-forrás hozzájárulásra utalnak44. Eredményeink rávilágítanak a lerakódási környezetek szélesebb körének vizsgálatának fontosságára, beleértve a megfelelő termikus érettségű és különböző litológiájú, szerves anyagban szegény rétegeket15,3034,59 , hogy pontosabb képet kapjunk a tengeri kémia és ökológia heterogenitásának helyenkénti nagyságrendjéről. Az óceánok fokozatos szellőzése, valamint a Rodinia felbomlása során és az ediakarán időszak alatt77,78 megnövekedett kémiai időjárás és tápanyagellátás ellenére az óceáni heterogenitás fenntartotta a különböző tengeri kémiai feltételeket, beleértve a tápanyagszegény, de lakható környezeteket, amelyek elősegítették a metazoák alkalmazkodását, versengését és evolúcióját a globális óceáni rendszerben (3. ábra). Míg az eutrofikus tengeri talapzatok demospongáknak adtak otthont, de gyakran hiányzott belőlük az Ediacara-biota, amire a dél-ománi sómedence (és az Ománban északabbra található korrelatív vastag Ediacara-kitermelések) kiváló példa; a sekély, oxikus és kevésbé produktív epikontinentális tengerszorosokat a demospongák helyett az Ediacara-biota népesítette be, annak ellenére, hogy a DOM és más szerves detrituszok helyben rendelkezésre álltak a táplálkozáshoz. Hogy a metabolikus követelmények vagy a környezeti szelekciós nyomás korlátozta-e az ediakari lágy testű többsejtű élővilágot ezekre a környezetekre, továbbra is bizonytalan; tanulmányunk azonban rávilágít arra, hogy az ediakari oligotróf környezetek potenciálisan döntő szerepet játszottak a makroszkopikus többsejtű szervezetek és a tengeri közösségek ökológiájának evolúciójában.