Poikkeuksellisen alhainen lämpökypsyysaste
Lämpökypsyys on tärkein yksittäinen tekijä, joka vaikuttaa muinaisten sedimenttien biomarkkereiden säilymiseen, sillä orgaaniset molekyylit ovat alttiita rakenteellisille ja stereokemiallisille muutoksille asteittaisen hautautumisen aikana ja lämpöstressin kasvaessa29. Prekambriset kivet, joiden lämpökypsyysaste on sopiva (öljyikkunan kypsyysaste tai sitä alhaisempi), ovat edellytys vankkojen ja primaaristen biomarkkerilipidiyhdistelmien säilymiselle. Useat hopaanin ja steraanin biomarkkerien stereoisomeerisuhteet sekä Rock-Eval-pyrolyysin alhaiset Tmax-arvot (enimmäkseen 417-433 °C:n välillä, keskiarvo 426 °C) osoittavat, että tämän tutkimuksen kivet edustavat ylivoimaisesti lämpökypsimpiä ediacaran-kiviä, joita on tähän mennessä analysoitu uusimmilla orgaanisen geokemian menetelmillä (taulukko 1). Vaikka kivemme ovat läpikäyneet sedimentaarisen diageneesin, ne eivät ole siirtyneet merkittävästi öljyikkunaan lukuun ottamatta Moldovan Podillyan altaasta peräisin olevia hieman kypsempiä 16PL-purkausnäytteitä, jotka soveltuvat edelleen analysoitaviksi (öljyikkunan alkuvaiheen tai keskivaiheen kypsyys, eikä niissä ole havaittavissa selviä merkkejä orgaanisista epäpuhtauksista). Suurin osa näytesarjasta ei todennäköisesti ollut altistunut yli 50 °C:n lämpötiloille, joten näytteemme ovat erittäin epäkypsiä. Muita molekulaarisia todisteita, jotka tukevat matalaa lämpökypsyyttä ja syngeenisyyttä, ovat i) polysyklisten biomarkkerialkaanien hallitsevuus n-alkaaneihin nähden kalliouutteissa (kuva 2), ii) havaittavissa olevien määrien 17β,21β(H)-hopaanien eloonjääminen, jotka voidaan erottaa runsaammin esiintyvistä hopaaneista, joilla on stabiilit 17β,21α(H)- ja 17α,21β(H)-stereokemialliset konfiguraatiot (taulukko 1, Täydentävät kuvat 1 ja 2). (taulukko 1 ja 2), (iii) havaittavissa oleva parittomien ja parillisten n-alkaanien välinen preferenssi C22-C27-alueella (kuva 2), koska n-alkaanit osoittavat hiililukupreferenssiä vain ennen katageneesiä38 , ja (iv) termisesti epäkypsien hopaanien ja steraanien biomarkkereiden tuottaminen (liukenemattomasta) kerogeenifaasista katalyyttisen hydropyrolyysin avulla (täydentävä kuva 2). Näytteidemme sedimenttisen orgaanisen aineksen alhainen terminen kypsyys vastaa aiemmin julkaistuja sedimenttimuutosindikaattoreita, kuten konodontti- ja akritarkki-muutosindeksejä, sekä aiempia Rock-Eval-pyrolyysi- ja biomarkkeritutkimuksia, jotka on tehty Ediacaran- ja paleotsooisista sedimenttikivistä Balticassa36,39,40 , sekä savimineralogiaa41. Kriittistä on, että ympäristöissä, joissa terminen kypsyys on näin alhainen, primääristen δ15N- ja δ13C-signaalien merkittävää muuttumista aiheuttavat mekanismit puuttuvat suurelta osin42.

Erottuvien alifaattisten hiilivetyjen jakauma edustavassa näytteessä. a Uuttuvien alifaattisten hiilivetyjen kokonaisionikromatogrammi (TIC) Lugovoe #13-73 m:lle Redkino Horizonista. N-alkaanisarja, pristaani (Pr), fytaani (Ph) ja C27-C34-hopaanit (merkitään niiden kokonaishiililuvun ja stereokemian mukaan C-17:ssä, C-21:ssä ja C-22:ssä, esim. C31αβR) on merkitty; ** merkitsee C29- ja C30-hopeenit. Huomaa, että hopaanien runsaus dominoi muita alkaaniyhdisteryhmiä. b Osittainen 85-Da-ionikromatogrammi osoittaa n-alkaanisarjan hallitsevuuden, jossa on havaittavissa parittomien ja parillisten hiililukujen välinen parittomien hiililukupreferenssi C22-C27-alueella, metyylialkaaneihin nähden
Lipidibiomarkkerikokoonpanojen mallit
Tärkeimpien (C27-C35) hopaanien ja tärkeimpien (C27-C29) steraanien runsauden suhdetta käytetään usein arvioitaessa bakteeri- ja eukaryoottisten lähdeorganismien tasapainoa muinaisessa vesiekosysteemissä. Hopaanit ovat molekyylifossiileja, jotka ovat peräisin hopanoideista, jotka ovat monenlaisten bakteeriryhmien syntetisoimia solukalvojen lipidejä. Vastaavasti steraanit ovat peräisin sterolien esiasteista, joita tuottavat lähes yksinomaan eukaryootit43. Näytteidemme hopaani/steraanisuhteet (H/St) kattavat hämmästyttävän laajan arvojen vaihteluvälin 1,6:sta 119,2:een (taulukko 1). Runsaasti orgaanisia aineita sisältävistä neoproterotsooisista kivistä ja öljyistä saadut H/St-suhteet ovat tyypillisesti kapealla välillä 0,5-2,030,31,32,33. Vaikka nuoremman Kotlin-horisontin (keskimääräinen H/St 8,9) ja vanhemman Redkino-horisontin (keskimääräinen H/St 42,9) arvot eroavat selvästi toisistaan, useimpien näytteiden arvot ovat epätavallisen korkeita ja viittaavat bakteerien poikkeuksellisen suureen osuuteen.
Eukaryoottiset levät olivat myöhäisellä neoproterotsooisella kaudella ekologisesti merkittävä komponentti ja päätuottajia monissa meriekosysteemeissä15,34,44. Epäjohdonmukaisuus eukaryoottisten levien globaalisti merkittävän osuuden välillä myöhäisellä neoproterotsooisella kaudella ja kuitenkin steraanibiomarkkereiden alhaiset tasot näissä näytteissä osoittavat, että eukaryoottisten levien runsauteen on oltava jokin paikallinen tekijä. Yksinkertaisin selitys näytteissämme (taulukko 1) havaituille erittäin korkeille hopaani/steraani-suhteille yhdessä matalien orgaanisen hiilen kokonaispitoisuuksien (TOC) ja matalien vetyindeksien (HI) kanssa on se, että nämä kerrostumat kerrostuivat oligotrofisiin (eli voimakkaasti ravinneköyhiin) olosuhteisiin, joissa bakteerit syrjäyttivät levät. Nykyaikaisia analogeja muinaisille epikratonisille merille, jotka kehittyivät korkean merenpinnan korkeuden aikana, on vaikea löytää, mutta samansuuntaisia havaintoja korkeammista (jopa suuruusluokkaa korkeammista) hopaani/steraanisuhteista on aiemmin havaittu orgaanisesti vähärasvaisten ja orgaanisesti runsasravinteisten sedimenttikivien osalta, jotka ovat kerrostuneet ordovikiumin ja siluurin aikaisiin epikontinentaalisiin meriväyliin45.
Tyypillisesti alhainen TOC-pitoisuus voi johtua orgaanisen aineksen rajallisesta laskeutumisesta matalan tuottavuuden olosuhteissa, orgaanisen aineksen vähäisestä säilymispotentiaalista vesipatsaassa tai sedimenteissä tai laimenemisesta suurella siliklastivirralla. Nykyaikaisissa valtamerissä tuottavuutta rajoittaa tavallisimmin välttämättömien ravinteiden (typpi, fosfori, pii ja rauta) alhainen määrä46 . Typpi, fosfori ja rauta ovat rajallisia avomerialueilla, joilla nousuvesi, pölypäästöt ja rannikon valumat eivät tuota riittäviä määriä ravinteita. Nykyaikaisissa oligotrofisissa ympäristöissä bakteerien ja eukaryoottisen biomassan suhde on suurempi kuin eutrofisissa tai mesotrofisissa ympäristöissä. Mielenkiintoista on, että Moldovan Redkino-horisontista peräisin olevat Podillya 16PL -löytönäytteet ovat fosforiittia sisältäviä mutakiviä, ja niistä saadaan huomattavasti alhaisemmat hopaani/steraanisuhteet (8,1:11,5) kuin muista taulukossa 1 esitetyistä Redkino-näytteistä (vaikkakin ne ovat silti huomattavasti korkeammat kuin ne, joita on löydetty Etelä-Omanin suola-altaassa tai orgaanisesti rikkaissa faneerotsooisissa sedimenteissä, jotka tyypillisesti kuuluvat kapeaan, 05-2,0), mikä saattaa viitata siihen, että fosfaatin lisääntynyt saatavuus on saattanut vaikuttaa suotuisasti paikallisesti havaittuun eukaryoottien ja bakteerien väliseen suhteeseen (kun otetaan huomioon, että typen isotooppisignatuurit ovat suurelta osin muuttumattomia, ks. seuraava jakso). Fosforin (P) pitoisuudet sekä P/Fetotal- ja P/Al-suhteet ovat muuten yleisesti ottaen alhaisia Podiljan, Ukrainan ja Viron47 myöhäisediacaran sedimenteissä, lukuun ottamatta tätä stratigrafista tasoa, jossa on fosforiittikyhmyjä. Vastaavasti Kotlinin ja Redkinon siliklastisten kivien P-pitoisuudet olivat alhaiset (~0,01 – ~0,1 painoprosenttia) Itä-Euroopan alustan koillisreunan kairaussydämessä11.
Baltikan epikontinentaalisten merien laaja ja matala pinnanmuodostus on saattanut ylläpitää fosforin tai muiden ravinteiden rajallisuutta reuna-alueilla, mikä johtuu fosfaatin autigeenisestä saostumisesta rautamineraalien kanssa oksisissa pintavesissä ja useiden alkuaineiden sitomisesta hyllysedimentteihin48. Jos fosfaatti oli rajoittava ravinne, biosaatavan fosforin lisääntyminen olisi lisännyt paikallista alkutuotantoa ja tarjonnut suotuisammat kasvuolosuhteet suurempikokoiselle yksisoluiselle kasviplanktonille49,50,51. Nykyaikaisessa valtamerijärjestelmässä merten pikosyanobakteerien, Prochlorococcus ja Synechococcus, tiedetään hallitsevan kasviplanktonin solujen lukumäärää ja biomassaa oligotrofisissa trooppisissa ja subtrooppisissa valtameriympäristöissä, mukaan luettuina fosfaattirajoitteiset oligotrofiset alueet valtameren pintavesissä52. Proklorokokki ja heterotrofinen SAR11 (Pelagibacter) kukoistavat monenlaisten sopeutumisten ansiosta, mukaan luettuina alhaiset energiakustannukset, jotka johtuvat pienistä genomeista ja alhaisista replikaatiovauhdeista, suuremmasta pinta-alan ja tilavuuden välisestä suhteesta pienempien solujen koon ansiosta sekä ylimääräisistä solunottotoiminnoista, joilla maksimoidaan ravinteiden hyödyntäminen53. Monet bakteerit pystyvät myös korvaamaan vähäisiä ravinteita, esim. hyödyntämällä sulfolipidejä fosforipitoisten lipidien sijasta fosforin puutteessa54 , tai käyttämällä vaihtoehtoisia substraatteja, esim. hankkimalla typpeä ilmakehän N2:sta typen sitomisen avulla55 , ravinnestressin lievittämiseksi. Tuore tutkimus meribakteeriplanktonin selviytymisestä oligotrofisissa ympäristöissä56 , joissa fosfaattia on niukasti saatavilla, viittaa siihen, että polyfosfaattiaineenvaihdunnalla on tärkeä rooli merten oligotrofisissa eliöissä. Valitut eukaryootit, jotka kilpailevat oligotrofisissa ympäristöissä, ovat tyypillisesti pieniä picoeukaryootteja, jotka saattavat täydentää ravinnontarvettaan mixotrofian avulla57. Picoeukaryoottien ja syanobakteerien biomassasuhde pyrkii kasvamaan, kun ravinteiden tarjonta lisääntyy49. Fosforin ja muiden biolimitoivien ravinteiden mahdollinen vaikutus primäärituottavuuden ja meriyhteisöjen rakenteen säätelyyn ediakarin epeiirisissä merialueilla vaatii lisätutkimuksia.
C29-steraanien runsaus on poikkeuksetta suurempi kuin vastaavien C27- tai C28-steraanien runsaus kaikissa paikoissamme (taulukko 1). C29-steraanien ylivoima C27- ja C28-steraaneihin nähden osoittaa todennäköisesti viherlevien hallitsevan eukaryoottista kasviplanktonyhteisöä44,58. Tämä piirre on havaittu useimmissa aiemmissa ediakaraanin biomarkkeritutkimuksissa15,30,31,32,33,34,35,36,44. Huomionarvoista on, että C30-steraanijakauma useissa näytteissä jokaisesta Kotlin-horisontin kairaussydämestä sisältää vähäisiä mutta havaittavissa olevia määriä 24-isopropyylikolestaanina (24-ipc)59,60 tunnettua demosponge-steraanibiomarkkeria. Kaikkiaan 24-ipc-steraaneja on raportoitu kivi- ja öljynäytteissä, jotka ovat peräisin Etelä-Omanin suola-altaasta niinkin kaukaa kuin kryogeenikaudelta (>635 Ma sitten), ja ne ovat vanhimmat lipidibiomarkkerit, jotka osoittavat metazoanien esiintymistä59,60. Näytteidemme 24-ipc-biomarkkerit olivat joko noin kertaluokkaa harvinaisempia (suhteessa C27-C30-steraanien kokonaismäärään ne olivat vain 0,06-0,61 %; keskiarvo = 0,22 %) verrattuna Etelä-Omanin ediacaran-kiviin ja -öljyihin (keskimäärin 1,7 %59 ) tai ne olivat havaitsemisrajan alapuolella, koska niiden runsaus oli suurimmassa osassa näytteistä häviävän pieni.
Typen ja orgaanisen hiilen isotooppisuhteet
Typpi-isotoopit voivat auttaa hahmottamaan typen kierron suhteellista tasapainoa ja sitä, missä määrin joko typen sitominen tai epätäydellinen denitrifikaatio olivat merkittäviä reittejä, jotka vaikuttivat meriyhteisöjen käytettävissä olevaan ravinnetaseeseen. Kun diatsotrofiset bakteerit sitovat molekulaarista typpeä vesipatsaan sitoutuneen typen puutteen vuoksi, sedimentin kokonaistypen isotooppiarvot voivat olla lähellä 0‰61. Tämä on vastakohta positiivisille typpi-isotooppisignaaleille (välillä +2 – +10‰), joiden moodi on +4 – +6‰ ja jotka on havaittu neoproterotsooisissa merisedimenteissä, jotka ovat kerrostuneet oletettavasti nitraattipitoisiin olosuhteisiin, joissa nitraatti on denitrifioitunut vain osittain62. Kaikkien näytteidemme typpi-isotooppiarvot yhtä poikkeavaa näytettä lukuun ottamatta (taulukko 1) kattavat rajallisen positiivisen vaihteluvälin +3,5 – +6,5‰, joka on päällekkäinen myöhäisen neoproterotsooisen ajan orgaanista ainetta sisältävien sedimenttikivien moodin kanssa62. Redox-analyysit viittaavat siihen, että vesipatsaassa vallitsivat hapelliset olosuhteet, joten tietojemme mukaan nitraatti hallitsi liuenneen epäorgaanisen typen määrää. δ15N-arvojen vaihteluväli viittaa myös siihen, että N2-fiksaatio ei ollut primäärituottajien ensisijainen typenhankintatapa. Pikemminkin typen kiertokulkua hallitsi todennäköisesti vesipatsaan kierrätys, ja orgaaninen typpi hapettui määrällisesti nitraatiksi, kuten nykyaikaisissa, läheisissä meriympäristöissä. Pohjaeläinten denitrifikaatio ja orgaanisen typen hautautuminen olisivat olleet liuenneen epäorgaanisen typen ensisijaiset nielut, ja δ15N-arvojen vaihteluväli viittaa siihen, että vesipatsaan epätäydellisellä denitrifikaatiolla, joka tyypillisesti johtaa huomattavaan 15N-rikastumiseen, oli vain vähäinen merkitys63. δ15N-arvojen vähäinen vaihtelu kuvastaa näin ollen todennäköisesti alustalle kulkeutuneen nitraatin δ15N:ää, ja vesipatsaan nitraatin vähäisen pelkistymisen ja N2-fiksaation vaikutus on vähäinen. Koska typen sitomisesta ei ole vahvaa δ15N-näyttöä, oletamme, että typpi ei ollut ensisijainen biologisesti rajoittava ravinne Baltian epikontinentaalisissa altaissa. Eristäytyminen jokien ja eolien fosforilähteistä sekä oksiset olosuhteet laajoissa, matalissa merissä sijaitsevissa epikontinentaalisissa altaissa ovat saattaneet tehostaa authigeenisen fosforiitin ja jälkimetallipitoisten faasien poistumista, mikä on johtanut ravinnerajoitukseen, joka on rajoittanut eukaryoottisten solujen kasvua ja tuotantoa.
Kokonaisorgaanisen hiilen isotooppiarvot (CTOC) vaihtelevat väliltä -23,0 – -33,9‰, ja suurimmat erot havaittiin eri kairauspisteiden välillä. Suhteellinen 13C-rikastuminen isotooppisuhteisiin, jotka ovat suurempia kuin n. -28‰, on ristiriidassa niiden tietojen kanssa, jotka on saatu Omanin eutrofisiin, avomerellisiin ympäristöihin kerrostuneista samanaikaisista kerrostumista30,31. Ilmoittamamme vaihteluväli on kuitenkin yleisesti ottaen johdonmukainen muiden samalta ajanjaksolta kerrostuneiden Baltian paikkojen δ13CTOC:n vaihteluvälin kanssa11. Ero Baltikan ja Omanin välillä saattaa korostaa sitä, että δ13CTOC-arvot eivät heijasta yksinomaan hiilen kiertokulun yhtenäistä sekulaarista muutosta tällä ajanjaksolla31,64. Pikemminkin Baltican δ13CTOC-arvojen vaihteluväli saattaa osittain heijastaa bakteerien hallitsemaa mikrobiologiaa, johon viittaavat ainutlaatuiset lipidien biomarkkerisuhteet. Pieni solukoko, suuri pinta-alan ja tilavuuden suhde ja hidas kasvunopeus oligotrofisissa olosuhteissa voivat lisätä fraktioitumisen suuruutta autotrofian aikana (εp)65,66 , mikä johtaa alhaisiin δ13C-arvoihin.
Mekanismi, joka johtaa 13C:llä rikastuneempiin isotooppisignaattoreihin myöhäisen ediakaraanin aikaisen Baltica-sukkession osalta raportoidun11 vaihteluvälien sisällä, ei ole yhtä selvä. Detritaalisen, metamorfisesti muuttuneen orgaanisen aineksen mahdollinen osuus prekambrikauden alhaisen TOC-pitoisuuden sedimenttisukkessioissa on mahdollinen mekanismi, joka selittää alueiden väliset erot δ13CTOC64 -arvoissa. Vaikka näytteidemme TOC-pitoisuus on alhainen, se on kuitenkin yleensä yli 0,10 painoprosenttia, mikä ei paljasta TOC-pitoisuuden ja δ13C-arvojen välistä yhteyttä. Lisäksi näytteissämme on termisesti kypsymätöntä orgaanista ainesta ja biomarkkerien lipidikuvioita, jotka ovat ristiriidassa pääasiassa alloktonisen hiililähteen kanssa. 13C-rikastuneemmat δ13CTOC-merkinnät voivat olla seurausta vaihtoehtoisista hiilen assimilaatiomekanismeista. Hiilikonsentraatiomekanismit tai prokaryoottien aktiivinen bikarbonaatin otto voivat johtaa pienempiin εp-arvoihin ja korkeampiin δ13CTOC-arvoihin67,68. Ediakaraanin eliöstön ilmaantuminen on saattanut laajentaa merkittävästi merten ravintoverkkoja ja edistää mikrobien heterotrofian uusia väyliä, mukaan luettuna monimutkaisen hiilenkierron mahdollinen vaikutus pohjaeläinten mikrobimattojen sisällä. Kun lisäksi otetaan huomioon liuenneen orgaanisen aineksen69 mahdollisesti tärkeä rooli hiilen lähteenä, δ13CTOC-arvojen laajuus kuvastaa todennäköisesti erilaisia biogeokemiallisia hiilen kierrätysprosesseja, jotka saattavat olla ainutlaatuisia kehittyvässä ediacaralaisessa meriympäristössä ja jotka saattavat liittyä ilmiöön, jolle meillä ei ole edustavia nykyaikaisia analogeja.
Etelä-Kiinan varhaisen triaskauden kivistä70 on löydetty muinaisia lipidibiomarkkerikokoonpanoja ja stabiilien isotooppien indikaattoreita, jotka viittaavat bentaaliseen, mikrobiperäiseen mattotuotantoon yhdessä kiinteän typen rajoituksen kanssa. Meishanissa tapahtui permikauden lopun massasukupuuton jälkimainingeissa suuri piikki humaani/steraani-suhteissa (noin 60:een asti), johon liittyi voimakas siirtymä Norgin isotooppisignaatiossa arvoihin 0-2‰, mikä on sopusoinnussa bakteerien diatsotrofian kanssa. Tähän liittyy 2-metyylihopaanien (2-metyylihopaani-indeksi jopa 33 %) ja omaleimaisten metyylialkaanien voimakas signaali, joka viittaa pohjaeläinten mikrobimattojen lisääntymiseen. Emme havaitse samanlaista suuntausta typpi-isotooppisignaaleissa tai biomarkkerikuvioissa, jotka ilmeisesti viittaisivat merkittävään mikrobimattojen syöttöön ediacaran-aineistossamme, vaikka näytteissämme havaittiin yksittäinen 15N-vähentynyt arvo (taulukko 1). Näin ollen Baltica-näytteissämme havaittu voimakas bakteerisignaali ei todennäköisesti ole pääasiassa bentaalisten mikrobimattojen merkki.
Paleoenvironmental sustenance of Ediacara biota vs. demosponges
Silminnähtävät oligotrofiset olosuhteet Baltican epikrateonisissa ja mannerlaidunalueiden altaissa, kun se ajautui korkeilta leveysasteilta matalille leveysasteille myöhäisestä ediakaaresta varhaiskambrikauden alkuun, saattavat liittyä joko suhteellisen ravinteikkaiden syvien vesien rajoitettuun advektioon tai ravinteiden köyhtymiseen, joka johtui assimilaatiosta ja haaskauksesta kuljetuksen ja laskeutumisen aikana näillä laajoilla, matalilla merialueilla sijaitsevilla, epikuriteollisilla vesistöalueilla (Fig. 3). Nämä altaat olivat ajoittain eristyksissä valtameristä, ja niissä kehittyi hypersuolapitoisuus (esim. Redkinon aikana) ja murtovesiolosuhteet (esim. Kotlinin aikana71). Pitkäaikainen tektoninen vakaus johti myöhäisen Ediacaran Baltican matalareliefiseen topografiaan, joka oli erittäin altis tulville ja sään vaikutuksesta peräisin olevan fosforin tehottomalle saannille. Toisin kuin Balticassa, biomarkkeritutkimukset Omanissa sijaitsevan Huqf-superryhmän kerrostumissa paljastivat eutrofisen ekosysteemin, jossa esiintyi runsaasti mikroleviä30,31,59 mutta ei Ediacara-biotooppia, jopa laajoissa esiintymissä Omanin vuoristossa ja Huqfin alueella, sisä- ja ulkohyllyjen välisissä ympäristöissä3. Etelä-Omanin suola-altaan paleogeografia on rekonstruoitu myöhäiselle neoproterotsooiselle ajalle ~13°:n etäisyydelle päiväntasaajasta eteläisellä pallonpuoliskolla, joka on suurin piirtein samankaltainen kuin Baltica72 -paleolaatikko.
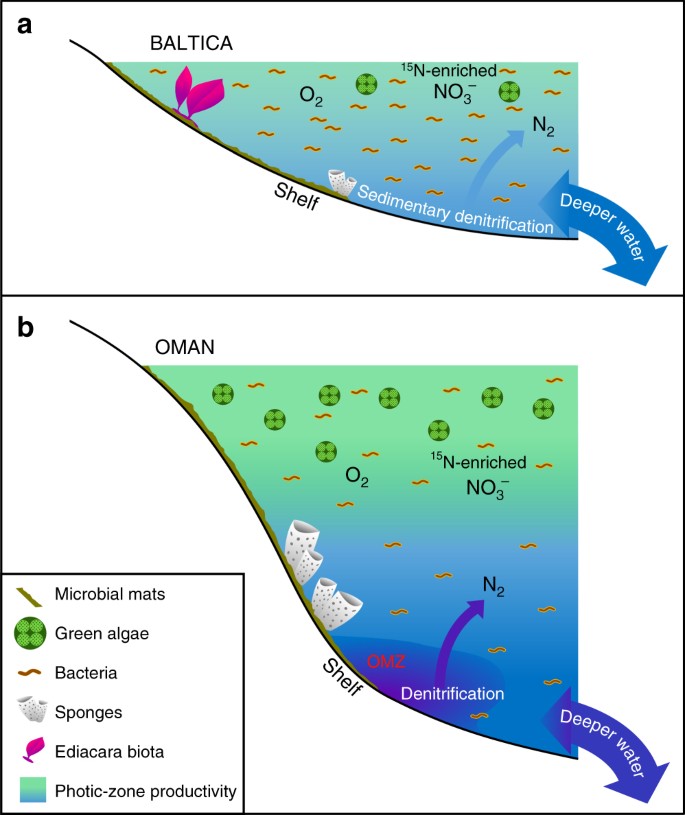
Merkittäviä eroavaisuuksia matalan tuottavuuden vs. tuottavien ediakarialaisten merellisten ympäristöjen välillä. Skemaattiset kaaviot on esitetty laajasta oligotrofisesta ja matalamerellisestä Baltian epikontinentaalisesta altaanreunasta, jota usein hallitsi bakteerituottavuus, jossa Ediacara-pehmyteläimistö kukoisti ja jossa denitrifikaatio ja anammox rajoittuivat todennäköisesti sedimentteihin; ja b Etelä-Omanin suola-altaan eutrofiset ja syvemmällä sijaitsevat syvänmeren hyllyalueet, joissa viherlevät kukoistivat primaarituottajina ja joissa oli runsaasti demospongeja, mutta Ediacara-pehmyteläimistö ei ollut merkittävä (Ediacara-eliöstön fossiileja ei ole myöskään Pohjois-Omanin vastaavissa Ediacaran-paljastumissa). Tuottavilla mannerreunoilla denitrifikaatiota ja anammoxia esiintyi todennäköisesti sekä vesipatsaassa että sedimenteissä
Pitkäkestoisesta ja laajamittaisesta anoksiasta Baltian matalilla mannerreunoilla ja niiden läheisyydessä myöhäisen ediakaraanin aikana ei ole todisteita11, ja ehdotamme, että tässä kuvatut oligotrofiset olosuhteet johtuivat maanpäällisen ja syvän veden tehottomasta ravinnevirrasta näihin ympäristöihin, mikä on pitkälti samanlaista kuin nykyisten valtamerijärjestelmien oligotrofisissa ympäristöissä. Pietarin alueen Utkina Zavodin ja sitä ympäröivien kairaussydänten geokemiallinen tutkimus viittaa siihen, että näytteemme kerrostuivat hapellisissa olosuhteissa73. Tätä tulkintaa tukevat itsenäisesti tietomme laajennetuista hopaanien (C31-C35) jakaumista, joiden runsaus vähenee jyrkästi C31-yhdisteiden yläpuolella hiililuvun kasvaessa, mikä on ominaista bakteriohopaanipolyolien sivuketjujen hajoamiselle diageneesin aikana oksisissa olosuhteissa29, ja myös alhaiset vetyindeksit, jotka on mitattu Rock-Eval-pyrolyysillä (kaikki alle 230 mg/g TOC, ja monet ovat alle 100 mg/g TOC) näille epäkypsille näytteille, jotka johtuvat uusiutuvan kerogeenin muodostumisesta hapettuvan hajoamisen ja lipidiköyhän primaaribiomassan uudelleenkondensoitumisen kautta paikallisesti oksisissa ympäristöissä (taulukko 1). Ediacaran monisoluisilla eliöillä, jotka elivät näissä matalissa meriympäristöissä, on täytynyt olla riittävästi orgaanista substraattia heterotrofiaa varten, jotta ne pystyivät tyydyttämään ravintotarpeensa, sekä riittävästi happea ja muita ravinteita aineenvaihdunnan ylläpitämiseksi. Baltican epikontinentaaliset altaat olivat todennäköisesti pysyvämmin hapekkaita kuin happiminimivyöhykkeitä reunustavat erittäin tuottavat ympäristöt, kuten Etelä-Omanin suola-altaan keski- ja ulkoselustalla, jossa runsaan planktisen biomassan hengitys olisi pitänyt liuenneen hapen alhaisemmalla tasolla fotisen vyöhykkeen alapuolella.
Ediacara- eliöstön ja muiden Baltican edustalla elävien monisoluisten eliöiden olisi täytynyt selviytyä vaihtelevista ravinnonlähteistä (bakteerit vs. eukaryoottiset), kun ravinnevirrat vaihtelivat ajan myötä, mukaan lukien pienet solut ja orgaaninen detritus epikontinentaalisissa altaissa, joissa bakteerit olivat hallitsevia primaarituottajia (kuva 3). Nykyaikaiset koralliriuttayhteisöt selviytyvät trooppisissa oligotrofisissa ympäristöissä tehokkaan ravinteiden kierrätyksen ansiosta, mukaan luettuna liuenneen orgaanisen aineen (DOM) virtaus, joka auttaa ylläpitämään eläimistön heterotrofiaa riuttaekosysteemissä74. Meren trofisen rakenteen, jossa eukaryoottiset monisoluiset organismit elävät orgaanisen detrituksen ravinnon avulla, on täytynyt syntyä vasta sen jälkeen, kun eukaryootit laajenivat globaalin mittakaavan ympäristölaajentumisen myötä erilaisiin meriympäristöihin, mikä tapahtui Tonian-Kryogenian-välin aikana (n. 800-635 Ma), kuten biomarkkeritietoaineistoista käy ilmi15. Tällöin voisimme odottaa havaitsevamme viitteitä merkittävistä eroista meriyhteisöjen ja trofian rakenteessa paikasta toiseen ediacaran-kauden loppupuolella, jolloin paikallinen ravinnetasapaino valitsi eukaryoottipitoisia tai eukaryoottipitoisia mikrobiyhteisöjä ja monisoluisten organismien asteittainen levittäytyminen lisäsi monimutkaisuuden toista ulottuvuutta eliö- ja yhteisötasolla. Merkittävät alueelliset erot fosfaatin ja muiden ravinteiden saatavuudessa hylly-ympäristöissä ovat myös odotettavissa oleva seuraus heterogeenisesta maailmanlaajuisesta merten redox-rakenteesta myöhäisellä ediakaarikaudella ennen syvän valtameren hapettumista38.
Pikoplanktonin bakteerituottavuuden ja siihen liittyvien liuenneen orgaanisen aineksen (DOM) hajoamistuotteiden hallitsevuus Baltican epikontinentaalisissa altaissa on voinut ylläpitää mikrobien kiertokulkuekosysteemiä rinnakkain perinteisen trofisen rakenteen kanssa, joka perustui suunnilleen suuremman kokoluokan planktonisiin tuottajiin50. Tämä on saattanut suosia erilaisia heterotrofian muotoja, kuten suspensioruokintaa ja mahdollisesti osmotrofiaa, joidenkin rangeomorfien, sienien ja muiden myöhäisediacaran aikakauden monisoluisten eliöiden18 käyttökelpoisena ravintostrategiana yhdessä kehittyvän aktiivisen (liikkuvan) heterotrofian kanssa19,20. Ravinneköyhiä vesijärjestelmiä hallitsevat yleensä usein pieni yksisoluinen kasviplankton ja heterotrofinen plankton, ja bakteerit kilpailevat eukaryoottien kanssa ja ylläpitävät vähäistä nettobiomassaa nykyisten valtamerten oligotrofisissa meriympäristöissä49,51. DOM on tärkeä orgaanisten ravinteiden lähde, ja se säätelee usein tuottavuutta ja nettobiomassaa nykyaikaisissa oligotrofisissa trooppisissa merissä, mutta niin on myös typen ja fosforin yhteisrajoitus75. Vastaavasti DOM oli todennäköisesti myös tärkeä substraatti heterotrofisten bakteerien ja mikrobikierron ylläpitämiselle muinaisissa oligotrofisissa ympäristöissä. Pohjan mikrobimatot ovat saattaneet olla osa tätä bakteerien hallitsemaa ravintoverkkoa, ja mattopohjien on todettu edistävän Ediacara-eliöstön säilymistä merialueilla7. Vaikka 2-metyyliopaani-indeksin kohtalaisia arvoja löytyi osajoukosta näytteistämme (4-10 %, taulukko 1), metyylialkaanien hyvin vähäiset määrät suhteessa n-alkaaneihin (kuva 1) ja karotenoidien vain vähäiset ja satunnaiset esiintymät viittaavat siihen, että mikrobimatot eivät hallinneet primäärituottavuutta76 ja että planktoniset bakteerit ja vastaavasti niiden DOM-jakautumistuotteet kukoistivat. Yleisesti ottaen alhaiset vetyindeksiarvot (taulukko 1; erityisesti Redkinon näytteiden osalta), huolimatta kerrostumien ja biomarkkerikokoonpanojen alhaisesta lämpökypsyydestä, ovat myös sopusoinnussa lipidiköyhän orgaanisen aineksen syötön kanssa pääasiassa hapellisissa, matalissa merissä sijaitsevissa marginaalisissa ympäristöissä. Mikrobimattojen esiintyminen pelkästään sedimentologisten rakenteiden perusteella ei missään tapauksessa kerro eukaryoottien (mikrolevien) ja bakteerien välisestä tasapainosta, eivätkä ne myöskään pysty määrittämään mikrobiplanktonin suhteellista osuutta yleiseen primaarituottavuuteen ja ravintoverkkojen ylläpitoon. Mikrobimattoyhteisöt, jotka sisältävät usein runsaasti eukaryootteja, ja Etelä-Omanin Ara-ryhmän karbonaatit, joissa on näkyvästi tromboliitti- ja ryppyisiä laminiittifasieita, ovat hyvä esimerkki myöhäisen neoproterotsooisen ajan ympäristöistä, joissa mikrobimattojen osuus on merkittävä, ja jotka tuottavat runsaasti steraanisignaaleja, jotka johtuvat mikrolevien suuresta osuudesta30,59.
24-ipc-biomarkkereiden erittäin alhainen määrä viittaa siihen, että vaikka demospongia esiintyi toisinaan näissä ympäristöissä vain vähän. Tämä saattaa viitata siihen, että pienemmillä suodattimia syövillä eläimillä on vähemmän mahdollisuuksia kilpailla Ediacaran eliöstön kanssa näissä matalan tuottavuuden ympäristöissä, demospongien riittämätön sietokyky energeettisempiä matalan meren olosuhteita vastaan reilun aallonpohjan yläpuolella tai sienien parempi sopeutumiskyky matalahappisiin olosuhteisiin, joita ylläpidetään dynaamisesti valovyöhykkeen alapuolella rehevöityneissä ympäristöissä (kuva 3). Kaiken kaikkiaan 24-ipc:tä ei havaittu useimmissa Redkino-horisontin näytteissä, joiden H/St-suhteet ovat yleensä myös näytejoukkomme korkeimmat, mikä saattaa viitata ekologiseen muutokseen Kotlin-horisontin kerrostumisajankohtaan mennessä. Vaihtoehtoisesti on myös uskottavaa, että sienet asuttivat näitä ympäristöjä, mutta eivät tuottaneet näitä diagnostisia steroidibiomarkkereita runsaasti, vaikka tämä vaikuttaa vähemmän todennäköiseltä, sillä 24-ipc-steraani on yksi yleisimmin havaituista C30-steraaniyhdisteistä ediacaran-kerrostumissa ja -öljyissä30,3133,59,60 , ja se on havaittavissa myös osajoukossa näytteistämme (taulukko 1).
Ja vaikka aineistomme viittaavat pysyviin oligotrofisiin meriympäristöihin, jotka edustavat paikallisia olosuhteita Ediacaran valtamerissä, ne eivät todennäköisesti olleet harvinaisia prekambrikauden matalien merialueiden merialueilla (kuva 3). Valtaosa aikaisemmista ediakaraanin biomarkkeritutkimuksista on tehty eutrofisiin ympäristöihin kerrostuneilla orgaanisesti rikkailla sedimenttikivillä ja niiden öljytuotteilla, jotka yleensä tuottavat biomarkkerikokoonpanoja, jotka ovat sopusoinnussa merkittävän mikrolevien osuuden kanssa44. Tuloksemme korostavat, että on tärkeää tutkia laajempaa valikoimaa laskeutumisympäristöjä, mukaan lukien orgaanisen aineen suhteen vähärasvaiset kerrostumat, joilla on sopiva terminen kypsyysaste ja erilaiset litologiat15,3034,59, jotta saadaan tarkempi kuva merikemian ja ekologian heterogeenisuuden laajuudesta paikasta toiseen. Huolimatta valtamerten asteittaisesta tuulettumisesta ja lisääntyneestä kemiallisesta säätilasta ja ravinteiden saannista Rodinian hajoamisen aikana ja koko Ediacaran-kauden ajan77,78 valtamerten heterogeenisuus säilytti erilaiset merikemialliset olosuhteet, mukaan luettuina ravinneköyhät mutta elinkelpoiset ympäristöt, jotka edistivät metazoan sopeutumista, kilpailua ja evoluutiota maailmanlaajuisessa valtameren järjestelmässä (kuva 3). Vaikka eutrofisilla merihyllyillä esiintyi demospongeja, Ediacara-eliöstö puuttui usein, mistä Etelä-Omanin suola-allas (ja siihen liittyvät paksut Ediacara-esiintymät pohjoisempana Omanissa) ovat näkyvä esimerkki; päinvastoin, matalat, hapekkaat ja vähätuottoisemmat epikontinentaaliset meriväylät asuttivat Ediacara-eliöstön mieluummin kuin demospongeja, huolimatta siitä, että DOM:aa (dityppioksidia) ja muuta orgaanista ainesta oli saatavissa ravinnoksi paikallisesti. Se, rajoittivatko aineenvaihdunnalliset vaatimukset tai ympäristön valikoivuuspaine Ediacaran pehmeärunkoiset monisoluiset eliöt näihin ympäristöihin, on edelleen epävarmaa; tutkimuksemme kuitenkin korostaa, että Ediacaran oligotrofisilla ympäristöillä oli potentiaalisesti ratkaiseva rooli makroskooppisten monisoluisten eliöiden evoluutiossa ja meriyhteisöjen ekologiassa.