Maturité thermique exceptionnellement faible. des strates
La maturité thermique est le facteur le plus important influençant la préservation des biomarqueurs sédimentaires anciens, car les molécules organiques sont susceptibles de subir une altération structurelle et stéréochimique au cours de l’enfouissement progressif et avec l’augmentation du stress thermique29. Les roches précambriennes de maturité thermique appropriée (maturité de la fenêtre pétrolière ou inférieure) sont des conditions préalables à la préservation d’assemblages lipidiques robustes et primaires de biomarqueurs. De multiples rapports de stéréoisomères de biomarqueurs hopane et stérane, ainsi que des preuves indépendantes de faibles valeurs de Tmax (principalement dans une plage de 417-433 °C, avec une moyenne de 426 °C) provenant de la pyrolyse Rock-Eval, indiquent que les roches de cette étude représentent de loin les roches édiacariennes les plus thermiquement immatures analysées à ce jour à l’aide des méthodes géochimiques organiques de pointe (tableau 1). Bien que nos roches aient subi une diagenèse sédimentaire, elles ne sont pas passées de manière significative dans la fenêtre pétrolière, à l’exception des échantillons d’affleurement 16PL légèrement plus matures du bassin de Podillya en Moldavie, qui sont encore adaptés à l’analyse (maturité de la fenêtre pétrolière précoce à moyenne et aucun signe évident de contaminants organiques). La majorité de l’ensemble n’avait alors probablement pas été exposée à des températures d’enfouissement supérieures à 50 °C, et nos échantillons sont donc très immatures. D’autres lignes de preuves moléculaires, qui soutiennent la faible maturité thermique et la syngénicité, comprennent (i) une dominance des alcanes biomarqueurs polycycliques par rapport aux n-alcanes dans les extraits de roche (Fig. 2), (ii) la survie de quantités détectables de 17β,21β(H)-hopanes résolubles à partir des hopanes plus abondants possédant des configurations stables 17β,21α(H)- et 17α,21β(H)-stéréochimiques (Tableau 1, Fig. supplémentaires. 1 et 2), (iii) une préférence discernable entre les impairs et les pairs parmi les n-alcanes dans la gamme C22 à C27 (Fig. 2), les n-alcanes montrant une préférence pour le nombre de carbones seulement avant la catagenèse38, et (iv) la génération de biomarqueurs hopane et stérane thermiquement immatures à partir de la phase kérogène (insoluble) en utilisant l’hydropyrolyse catalytique (Fig. 2 supplémentaire). La faible maturité thermique de la matière organique sédimentaire dans nos échantillons est cohérente avec les indicateurs d’altération sédimentaire précédemment publiés, y compris les indices d’altération des conodontes et des acritarches, et les études précédentes de pyrolyse et de biomarqueurs Rock-Eval des roches sédimentaires édiacariennes et paléozoïques de Baltica36,39,40, ainsi que la minéralogie des argiles41. De manière critique, dans les milieux où la maturité thermique est aussi faible, les mécanismes d’altération significative des signaux primaires δ15N et δ13C sont largement absents42.

Distribution des hydrocarbures aliphatiques extractibles pour un échantillon représentatif. a Chromatogramme d’ions totaux (TIC) pour les hydrocarbures aliphatiques extractibles pour Lugovoe #13-73 m de l’horizon Redkino. La série des n-alcanes, le pristane (Pr), le phytane (Ph) et les hopanes C27-C34 (désignés par leur nombre total de carbone et leur stéréochimie en C-17, C-21 et C-22, par exemple C31αβR) sont étiquetés ; ** désigne les hopènes C29 et C30. Notez la dominance de l’abondance des hopènes par rapport aux autres classes de composés d’alcane. b Le chromatogramme partiel d’ions 85-Da montre la prédominance de la série des n-alcanes, avec une préférence discernable pour les nombres impairs ou pairs de carbone dans la gamme C22-C27, sur les méthylalcanes
Modèles d’assemblage de biomarqueurs lipidiques
Un rapport d’abondance des principaux hopanes (C27-C35) sur les principaux stéroanes (C27-C29) est souvent utilisé pour évaluer l’équilibre des apports d’organismes sources bactériens par rapport aux eucaryotes dans l’ancien écosystème aquatique. Les hopanes sont des fossiles moléculaires dérivés des hopanoïdes, qui sont des lipides de membrane cellulaire synthétisés par une grande variété de groupes bactériens. De même, les stérols sont dérivés de précurseurs de stérols, qui sont produits presque exclusivement par les eucaryotes43. Les rapports hopane/stérane (H/St) de nos échantillons couvrent une gamme étonnamment large de valeurs allant de 1,6 à 119,2 (Tableau 1). À titre de comparaison, les rapports H/St des roches et huiles néoprotérozoïques riches en matières organiques se situent généralement dans une fourchette étroite allant de 0,5 à 2,030,31,32,33. Bien qu’il y ait une nette différence entre les valeurs de l’horizon Kotlin plus jeune (H/St moyen de 8,9) et celles de l’horizon Redkino plus ancien (H/St moyen de 42,9), les valeurs de la plupart des échantillons sont inhabituellement élevées et suggèrent des contributions anormalement élevées de bactéries.
À la fin du Néoprotérozoïque, les algues eucaryotes étaient une composante écologiquement significative et des producteurs majeurs dans de nombreux écosystèmes marins15,34,44. La divergence entre la contribution globalement importante des algues eucaryotes à la fin du Néoprotérozoïque et les faibles niveaux de biomarqueurs de stéroïdes dans ces échantillons indique qu’il doit y avoir un déterminant local sur l’abondance des eucaryotes. L’explication la plus parcimonieuse des rapports hopane/stérane extrêmement élevés, des faibles teneurs en carbone organique total (COT) et des faibles indices d’hydrogène (IH) trouvés dans nos échantillons (tableau 1) est que ces strates ont été déposées dans des milieux oligotrophes (c’est-à-dire fortement limités en nutriments), dans lesquels les bactéries ont supplanté les algues. Bien qu’il soit difficile de trouver des analogues modernes pour les anciennes mers épicratoniques qui se sont développées pendant un haut niveau de la mer, des observations parallèles de rapports hopane/stérane plus élevés (jusqu’à un ordre de grandeur) ont été trouvées précédemment pour les roches sédimentaires pauvres en matières organiques par rapport aux roches sédimentaires riches en matières organiques déposées dans les voies maritimes épicontinentales de l’Ordovicien-Silurien45.
Typiquement, une faible teneur en COT peut être causée par un dépôt limité de matière organique dans des milieux à faible productivité, un faible potentiel de conservation de la matière organique dans la colonne d’eau ou les sédiments, ou une dilution avec un flux siliciclastique élevé. Dans les océans modernes, la productivité est le plus souvent limitée par de faibles niveaux de nutriments essentiels : azote, phosphore, silicium et fer46. L’azote, le phosphore et le fer sont limités dans les régions de haute mer où les remontées d’eau, l’apport de poussière et le ruissellement côtier ne fournissent pas des quantités suffisantes de nutriments. Dans les milieux oligotrophes modernes, le rapport entre la biomasse bactérienne et la biomasse eucaryote est plus élevé que dans les milieux eutrophes ou mésotrophes. Il est intéressant de noter que les échantillons de l’affleurement Podillya 16PL de l’horizon Redkino de Moldavie sont des mudstones contenant de la phosphorite et donnent des rapports hopane/stérane significativement plus faibles (8,1:11,5) que les autres échantillons Redkino du tableau 1 (bien que toujours significativement plus élevés que ceux trouvés dans le bassin salin d’Oman Sud ou dans les sédiments phanérozoïques riches en matières organiques, qui se situent généralement dans une gamme étroite de 0.5-2,0), ce qui pourrait suggérer qu’une disponibilité accrue de phosphate aurait pu influencer favorablement le rapport eucaryotes/bactéries trouvé localement (étant donné que les signatures isotopiques de l’azote sont largement invariables, voir la section suivante). Les teneurs en phosphore (P), ainsi que les rapports P/Fetotal et P/Al, sont par ailleurs généralement faibles pour les sédiments de l’Édiacarien supérieur de Podillya, d’Ukraine et d’Estonie47 ; à l’exception de ce niveau stratigraphique marqué par des nodules de phosphorite. De même, des niveaux faibles (~0,01 à ~0,1 % en poids) de P dans les roches siliciclastiques de Kotlin et Redkino ont été signalés dans une carotte de forage de la marge nord-est de la plate-forme est-européenne11.
La topographie large et peu profonde des mers épicontinentales à travers la Baltique pourrait avoir soutenu les limitations de phosphore ou d’autres nutriments dans les milieux marginaux, en raison de la précipitation autochtone du phosphate avec des minéraux de fer dans les eaux de surface oxiques et de la séquestration d’une gamme d’éléments par les sédiments du plateau continental48. Si le phosphate était un nutriment limitant, l’augmentation du phosphore biodisponible aurait amélioré la production primaire locale et fourni des conditions de croissance plus favorables pour le phytoplancton unicellulaire de plus grande taille49,50,51. En ce qui concerne le système océanique moderne, il est reconnu que les picocyanobactéries marines, Prochlorococcus et Synechococcus, dominent le nombre de cellules et la biomasse du phytoplancton dans les milieux océaniques oligotrophes tropicaux et subtropicaux, y compris les régions oligotrophes limitées en phosphate des eaux de surface océaniques52. Prochlorococcus et le SAR11 hétérotrophe (Pelagibacter) prospèrent grâce à une variété d’adaptations, y compris de faibles coûts énergétiques en vertu de petits génomes et de faibles taux de réplication, un rapport surface/volume plus élevé grâce à des cellules de plus petite taille, et des fonctions supplémentaires d’absorption cellulaire pour maximiser l’utilisation des nutriments53. De nombreuses bactéries sont également capables de substituer des nutriments peu abondants, par exemple en utilisant des sulfolipides au lieu de lipides phosphorés dans des environnements déficients en P54, ou en utilisant des substrats alternatifs, par exemple en s’approvisionnant en azote à partir du N2 atmosphérique via la fixation de l’azote55, pour atténuer le stress nutritionnel. Une étude récente concernant la survie du bactérioplancton marin dans des environnements oligotrophes56, où le phosphate disponible est limité, suggère un rôle important pour le métabolisme des polyphosphates chez les oligotrophes marins. Les quelques eucaryotes qui entrent en compétition dans les milieux oligotrophes sont généralement de petits picoeucaryotes, qui peuvent compléter leurs besoins nutritionnels par la mixotrophie57. Le rapport entre la biomasse des picoeucaryotes et celle des cyanobactéries a tendance à augmenter en cas d’apport accru de nutriments49. L’influence possible du phosphore, et d’autres nutriments biolimitants, dans la modération de la productivité primaire et de la structure des communautés marines dans les voies maritimes épeiriques de l’Édiacarien nécessite une étude plus approfondie.
Sans exception, l’abondance des stérols C29 est supérieure aux stérols C27 ou C28 correspondants pour tous nos sites (Tableau 1). Une prédominance des stérols C29 par rapport aux stérols C27 et C28 indique probablement une dominance des algues vertes au sein de la communauté phytoplanctonique eucaryote44,58. Cette caractéristique a été observée dans la plupart des études précédentes sur les biomarqueurs de l’Édiacarien15,30,31,32,33,34,35,36,44. Notamment, la distribution de stéroïdes C30 dans plusieurs échantillons de chaque carotte de forage dans l’horizon Kotlin contient des quantités faibles, mais détectables, du biomarqueur de stéroïdes de démosponge connu sous le nom de 24-isopropylcholestane (24-ipc)59,60. Au total, des stéroïdes 24-ipc ont été signalés dans des échantillons de roche et de pétrole datant d’aussi loin que le Cryogénien (>635 Ma) dans le bassin salin d’Oman Sud et représentent les plus anciennes preuves de biomarqueurs lipidiques pour les métazoaires59,60. Les biomarqueurs 24-ipc dans nos échantillons étaient soit d’un ordre de grandeur inférieur en abondance (par rapport aux ratios totaux de stéroïdes C27 à C30, ceux-ci n’étaient que de 0,06 à 0,61 % ; moyenne = 0,22 %) par rapport aux roches et huiles édiacariennes du sud d’Oman (1,7 % en moyenne59), soit inférieurs aux limites de détection en raison d’une abondance négligeable pour la majorité des échantillons.
Rapports isotopiques de l’azote et du carbone organique
Les isotopes de l’azote peuvent aider à discerner l’équilibre relatif du cycle de l’azote, et le degré auquel la fixation de l’azote ou la dénitrification incomplète étaient des voies significatives pour influencer l’équilibre des nutriments disponibles pour les communautés marines. Lorsque les bactéries diazotrophes fixent l’azote moléculaire en raison d’un manque d’azote fixé dans la colonne d’eau, cela peut donner des valeurs isotopiques de l’azote global sédimentaire proches de 0‰61. Cela contraste avec les signatures isotopiques positives de l’azote (dans la gamme de +2 à +10‰) avec un mode de +4 à +6‰ trouvé pour les sédiments marins néoprotérozoïques déposés dans ce que l’on pense être des conditions de nitrate-replet où le nitrate n’a subi qu’une dénitrification partielle62. Les valeurs isotopiques de l’azote pour tous nos échantillons, à l’exception d’une valeur aberrante (tableau 1), couvrent une gamme positive limitée de +3,5 à +6,5‰, qui recouvre le mode pour les roches sédimentaires riches en matière organique de la fin du Néoprotérozoïque62. Contraintes par les proxies redox suggérant que les conditions de la colonne d’eau oxique ont prévalu, nos données impliquent que le nitrate a dominé le pool d’azote inorganique dissous. La gamme des valeurs δ15N suggère également que la fixation de N2 n’était pas le principal mode d’acquisition d’azote pour les producteurs primaires. Le cycle de l’azote était plutôt dominé par le recyclage de la colonne d’eau, et l’azote organique était sujet à une oxydation quantitative en nitrate, comme c’est le cas dans les milieux marins modernes et proximaux. La dénitrification benthique et l’enfouissement de l’azote organique auraient été les principaux puits pour l’azote inorganique dissous, et la gamme des valeurs δ15N suggère seulement un rôle limité pour la dénitrification incomplète de la colonne d’eau, qui entraîne généralement un enrichissement significatif en 15N63. La variabilité limitée des valeurs δ15N reflète donc probablement le δ15N du nitrate advecté sur la plate-forme, avec une influence mineure de la réduction limitée du nitrate de la colonne d’eau et de la fixation de N2. Sans une forte évidence δ15N pour la fixation de l’azote, nous supposons que l’azote n’était pas le principal nutriment biolimitant dans les bassins épicontinentaux de Baltica. L’isolement des sources fluviales et éoliennes de phosphore, et les conditions oxiques dans les larges bassins épicontinentaux à faible profondeur peuvent avoir favorisé l’élimination de la phosphorite authigène et des phases contenant des métaux-traces, ce qui a entraîné une limitation des nutriments qui a contraint la croissance et la production des cellules eucaryotes.
Les valeurs isotopiques du carbone organique total (CTOC) varient de -23,0 à -33,9‰, les plus grandes différences étant observées entre les différents emplacements des carottes de forage. L’enrichissement relatif en 13C des rapports isotopiques supérieurs à environ -28‰ contraste avec les données provenant de strates contemporaines déposées dans les milieux marins ouverts eutrophes d’Oman30,31. Cependant, la gamme que nous rapportons est généralement cohérente avec la gamme δ13CTOC pour d’autres endroits de Baltica déposés au cours de la même période11. La différence entre Baltica et Oman pourrait souligner le fait que les valeurs δ13CTOC ne reflètent pas exclusivement un changement séculaire uniforme dans le cycle du carbone pendant cet intervalle de temps31,64. Plutôt, la gamme δ13CTOC de Baltica peut, en partie, refléter l’écologie microbienne dominée par les bactéries, suggérée par les rapports uniques de biomarqueurs lipidiques. La petite taille des cellules, les rapports surface/volume élevés, et les taux de croissance lents dans des conditions oligotrophes peuvent augmenter l’ampleur du fractionnement pendant l’autotrophie (εp)65,66, ce qui entraîne de faibles valeurs de δ13C.
Le mécanisme pour les signatures isotopiques plus enrichies en 13C dans la gamme rapportée11 pour la succession de Baltica de l’Édiacarien tardif est moins clair. La contribution potentielle de la matière organique détritique, altérée par métamorphisme, aux successions sédimentaires précambriennes à faible TOC est un mécanisme possible pour les disparités entre les sites en δ13CTOC64. Cependant, la teneur en COT de nos échantillons, bien que faible, est généralement supérieure à 0,10 % en poids, ce qui ne révèle aucune relation entre la teneur en COT et les valeurs de δ13C. De plus, nos échantillons contiennent de la matière organique thermiquement immature et des modèles de lipides biomarqueurs qui sont incompatibles avec une source de carbone principalement allochtone. Des signatures δ13CTOC plus enrichies en 13C peuvent être le résultat de mécanismes alternatifs d’assimilation du carbone. Les mécanismes de concentration du carbone ou l’absorption active de bicarbonate par les procaryotes peuvent entraîner des valeurs plus petites pour εp et des valeurs plus élevées de δ13CTOC67,68. L’émergence du biote édiacarien peut avoir considérablement élargi les réseaux alimentaires marins et stimulé de nouvelles voies d’hétérotrophie microbienne, y compris les contributions possibles du cycle complexe du carbone au sein des tapis microbiens benthiques. Avec une considération supplémentaire pour le rôle potentiellement important de la matière organique dissoute69 en tant que source de carbone, l’ampleur des valeurs δ13CTOC reflète probablement une gamme de processus biogéochimiques de cycle du carbone qui pourraient être uniques à l’environnement marin édiacarien en évolution et pourraient être liés à un phénomène pour lequel nous n’avons pas d’analogues modernes représentatifs.
Des assemblages de biomarqueurs lipidiques anciens et des indicateurs à isotopes stables pour la production de tapis microbiens benthiques en tandem avec une limitation de l’azote fixe ont été trouvés dans les roches du Trias précoce de Chine du Sud70. À la suite de l’extinction massive de la fin du Permien à Meishan, il y a eu un pic important dans les rapports hopane/stérane (jusqu’à environ 60) accompagné d’un fort changement dans la signature isotopique de Norg vers des valeurs de 0 à -2‰, ce qui est cohérent avec la diazotrophie bactérienne. Ceci est associé à un signal élevé de 2-méthylhopanes (indice 2-méthylhopane jusqu’à 33%) et de méthylalcanes distinctifs qui indiquent la prolifération de tapis microbiens benthiques. Nous n’observons pas de tendance similaire dans la signature isotopique de l’azote ou dans les modèles de biomarqueurs qui indiqueraient manifestement un apport significatif de tapis microbien dans notre ensemble de données édiacariennes, bien qu’une valeur singulière appauvrie en 15N ait été trouvée dans nos échantillons (Tableau 1). Par conséquent, le fort signal bactérien observé dans nos échantillons de Baltica n’est probablement pas principalement une signature des tapis microbiens benthiques.
Subsistance paléoenvironnementale du biote d’Ediacara vs. demosponges
Les conditions oligotrophes apparentes à travers les bassins épicratoniques et de marge continentale de Baltica, alors qu’elle dérivait des hautes aux basses latitudes de la fin de l’Édiacarien au début du Cambrien, pourraient être associées soit à une advection limitée d’eaux profondes relativement riches en nutriments, soit à un appauvrissement en nutriments, résultant de l’assimilation et du piégeage pendant le transport et le dépôt à travers ces larges bassins épicontinentaux à faible profondeur (Fig. 3). Ces bassins ont été épisodiquement isolés des océans et ont développé une hypersalinité (par exemple, à l’époque Redkino) et des conditions saumâtres (par exemple, à l’époque Kotlin71). La stabilité tectonique à long terme a donné lieu à une topographie à faible relief de la Baltica de l’Édiacara tardif, très sensible aux inondations et à un approvisionnement inefficace en phosphore dérivé de l’altération. Contrairement à Baltica, les études de biomarqueurs des strates du Supergroupe de Huqf à Oman ont révélé un écosystème eutrophique, riche en microalgues30,31,59, mais dépourvu de biote d’Ediacara, même dans les affleurements étendus des Monts Oman et de la région de Huqf, dans les milieux du plateau interne et externe3. La paléogéographie du bassin salé d’Oman Sud a été reconstituée pour la fin du Néoprotérozoïque à ~13° de l’équateur dans l’hémisphère sud, largement similaire à la paléolatitude Baltica72.
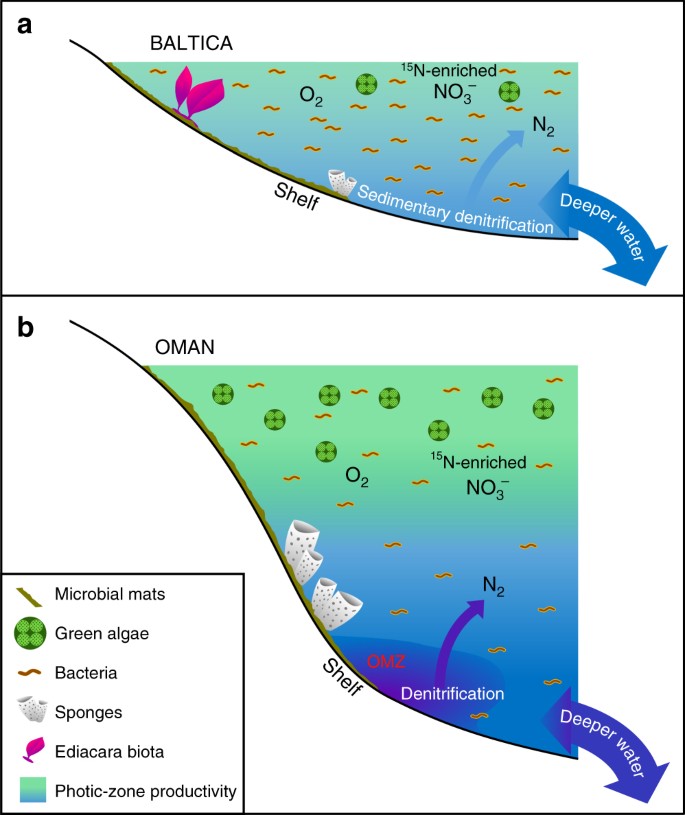
Différences majeures dans les environnements marins édiacariens peu productifs vs productifs. Des diagrammes schématiques sont présentés pour a des marges de bassin épicontinentales oligotrophes et peu profondes de la Baltique souvent dominées par la productivité bactérienne où la faune à corps mou d’Ediacara a prospéré et où la dénitrification et l’anammox étaient probablement limitées aux sédiments ; et b les milieux eutrophiques et les plateaux marins plus profonds du bassin salé du sud d’Oman, où les algues vertes ont prospéré en tant que producteur primaire et où les démosponges étaient abondantes, mais où la faune à corps mou d’Ediacara n’était pas proéminente (les fossiles de biote d’Ediacara étant également absents des affleurements d’Ediacara corrélatifs dans le nord d’Oman). Sur les marges continentales productives, la dénitrification et l’anammox se sont probablement produites à la fois dans la colonne d’eau et les sédiments
Il n’y a aucune preuve d’une anoxie persistante et étendue sur et autour des marges continentales peu profondes de Baltica pendant l’Édiacarien tardif11, et nous suggérons que les conditions oligotrophes décrites ici ont été causées par des flux inefficaces de nutriments terrestres et d’eau profonde vers ces milieux, largement similaires à ceux des environnements oligotrophes des systèmes océaniques modernes. En effet, une étude géochimique des éléments traces de l’Utkina Zavod et des carottes de forage adjacentes de la région de Saint-Pétersbourg suggère que nos échantillons ont été déposés dans des conditions oxiques73. Cette interprétation est indépendamment soutenue par nos données des distributions étendues d’hopane (C31-C35) qui se terminent brusquement dans l’abondance au-dessus des composés C31 avec l’augmentation du nombre de carbone, ce qui est caractéristique de la dégradation de la chaîne latérale des bacteriohopanepolyols pendant la diagenèse dans des conditions oxiques29, et également par des indices de faible teneur en hydrogène, comme mesuré par la pyrolyse Rock-Eval (tous inférieurs à 230 mg/g TOC, et beaucoup sont inférieurs à 100 mg/g TOC) pour ces échantillons immatures en raison de la formation de kérogène récalcitrant via la dégradation oxydative et la recondensation de la biomasse primaire pauvre en lipides dans des environnements localement oxiques (Tableau 1). Les organismes multicellulaires de l’Édiacara qui existaient dans ces environnements marins peu profonds devaient disposer de suffisamment de substrats organiques pour l’hétérotrophie afin de satisfaire leurs besoins alimentaires, ainsi que de suffisamment d’oxygène et d’autres nutriments pour soutenir leur métabolisme. Les bassins épicontinentaux de Baltica étaient probablement plus durablement oxiques que les milieux hautement productifs qui frangeaient les zones de minimum d’oxygène, comme sur le plateau moyen à extérieur du bassin salé d’Oman Sud, où la respiration de l’abondante biomasse planctonique aurait maintenu un oxygène dissous plus faible sous la zone photique.
Le biote d’Ediacara et d’autres organismes multicellulaires vivant au large de Baltica auraient dû faire face à des sources de nourriture changeantes (bactériennes vs eucaryotes) alors que les flux de nutriments variaient dans le temps, y compris les petites cellules et les détritus organiques dans les bassins épicontinentaux où les bactéries étaient les producteurs primaires dominants (Fig. 3). Les communautés modernes de récifs coralliens survivent dans les milieux oligotrophes tropicaux grâce à un recyclage efficace des nutriments, y compris la génération d’un flux de matière organique dissoute (DOM) qui contribue à soutenir l’hétérotrophie de la faune au sein de l’écosystème récifal74. L’établissement d’une structure trophique marine avec des organismes multicellulaires eucaryotes se nourrissant de détritus organiques a dû être postérieur à l’expansion environnementale à l’échelle mondiale des eucaryotes dans divers environnements marins, qui s’est produite pendant l’intervalle Tonien-Cryogène (environ 800-635 Ma), comme le montrent les enregistrements de biomarqueurs15. Nous pourrions alors nous attendre à discerner des preuves de différences significatives dans la communauté marine et la structure trophique d’une localité à l’autre au cours de la période de l’Édiacarien tardif, l’équilibre nutritif local sélectionnant des communautés microbiennes riches en eucaryotes ou pauvres en eucaryotes et l’expansion progressive des organismes multicellulaires ajoutant une autre dimension de complexité au niveau de l’organisme et de la communauté. Un contraste régional significatif dans la disponibilité du phosphate et d’autres nutriments dans les environnements de plateau est également une conséquence attendue d’une structure redox marine globale hétérogène pour la fin de l’Édiacarien, avant l’oxygénation de l’océan profond38.
La dominance de la productivité bactérienne du picoplancton et des produits de dégradation de la matière organique dissoute (DOM) associés dans les bassins épicontinentaux de la Baltique pourrait avoir soutenu un écosystème de boucle microbienne en parallèle avec la structure trophique conventionnelle basée sur autour des producteurs planctoniques de plus grande taille50. Cela peut avoir favorisé différents modes d’hétérotrophie, y compris l’alimentation en suspension et, peut-être, l’osmotrophie, comme stratégie d’alimentation viable pour certains rangeomorphes, éponges et autres organismes multicellulaires de la fin de l’Édiacarien18, en conjonction avec l’hétérotrophie active (mobile) émergente19,20. En règle générale, les systèmes aquatiques limités en nutriments sont souvent dominés par un petit phytoplancton unicellulaire et un plancton hétérotrophe, les bactéries surpassant les eucaryotes et maintenant une faible biomasse nette dans les milieux marins oligotrophes de l’océan moderne49,51. La DOM est une source importante de nutriments organiques et contrôle souvent la productivité et la biomasse nette dans les mers tropicales oligotrophes modernes, mais il en va de même pour la co-limitation de l’azote et du phosphore75. De même, la DOM était probablement un substrat important pour le maintien des bactéries hétérotrophes et une boucle microbienne dans les anciens milieux oligotrophes. Les tapis microbiens benthiques peuvent avoir été une composante de ce réseau alimentaire dominé par les bactéries, et les sols des tapis ont été impliqués dans l’amélioration de la préservation du biote d’Ediacara dans les milieux marins7. Alors que des valeurs modérées de l’indice de 2-méthylhopane ont été trouvées pour un sous-ensemble de nos échantillons (4-10%, Tableau 1), les très faibles abondances de méthylalcanes par rapport aux n-alcanes (Fig. 1), et seulement des traces et des occurrences sporadiques de caroténoïdes suggèrent que les tapis microbiens n’ont pas dominé la productivité primaire76, et que les bactéries planctoniques et, en conséquence, leurs produits de décomposition DOM ont prospéré. Les valeurs généralement faibles de l’indice d’hydrogène (tableau 1 ; en particulier pour les échantillons de Redkino), malgré la faible maturité thermique des strates et des assemblages de biomarqueurs, sont également cohérentes avec un apport organique pauvre en lipides dans des milieux marginaux marins peu profonds et principalement oxiques. Les occurrences de tapis microbiens, évaluées uniquement à partir des textures sédimentologiques, ne nous renseignent en aucun cas sur l’équilibre entre les eucaryotes (microalgues) et les bactéries, et ne peuvent pas non plus déterminer la contribution relative du plancton microbien à la productivité primaire globale et à la subsistance des réseaux alimentaires. Les communautés de tapis microbiens qui contiennent souvent des eucaryotes abondants et les carbonates du groupe Ara du sud d’Oman avec des faciès de laminite thrombolitique et frisée proéminents sont un bon exemple d’environnements de la fin du Néoprotérozoïque avec une contribution significative de tapis microbiens, donnant des signaux de stérane abondants en raison d’une grande contribution de microalgues30,59.
Le niveau extrêmement faible de biomarqueurs 24-ipc suggère que si les démosponges étaient parfois présents, ils étaient rares dans ces environnements. Cela pourrait indiquer moins d’opportunités pour les petits animaux filtreurs en compétition avec le biote d’Ediacara dans ces milieux à faible productivité, une résilience insuffisante des démosponges face aux conditions marines peu profondes plus énergétiques au-dessus de la base des vagues de beau temps, ou une meilleure adaptabilité des éponges aux conditions de faible teneur en oxygène maintenues dynamiquement sous la zone photique dans les milieux eutrophes (Fig. 3). Au total, le 24-ipc n’a pas été détecté dans la plupart des échantillons de l’horizon Redkino, qui ont aussi généralement les rapports H/St les plus élevés parmi notre ensemble d’échantillons, ce qui indique peut-être un changement écologique au moment du dépôt de l’horizon Kotlin. Alternativement, il est également plausible que les éponges aient habité ces milieux, mais n’aient pas produit ces biomarqueurs stéroïdes diagnostiques en abondance, bien que cela semble moins probable, car le 24-ipc stérane fait partie des composés stéroïdes C30 les plus couramment détectés dans les strates et les huiles édiacariennes30,3133,59,60 et est également détectable dans un sous-ensemble de nos échantillons (tableau 1).
Bien que les environnements marins oligotrophes persistants suggérés par nos données représentent des conditions localisées dans les océans édiacariens, ils n’étaient probablement pas rares pour les voies maritimes précambriennes à faible profondeur (Fig. 3). La grande majorité des études précédentes sur les biomarqueurs de l’Édiacarien ont été menées sur des roches sédimentaires riches en matières organiques déposées dans des milieux eutrophes et sur leurs produits pétroliers, qui produisent généralement des assemblages de biomarqueurs compatibles avec une contribution importante des microalgues44. Nos résultats soulignent l’importance d’étudier une plus grande variété d’environnements de dépôt, y compris des strates pauvres en matière organique de maturité thermique appropriée et de différentes lithologies15,3034,59, afin d’obtenir une image plus précise de l’échelle d’hétérogénéité de la chimie et de l’écologie marines d’un endroit à l’autre. Malgré la ventilation progressive de l’océan et l’augmentation de l’altération chimique et de l’apport en nutriments pendant la dislocation de Rodinia et tout au long de la période édiacarienne77,78, l’hétérogénéité océanique a maintenu une variété de conditions chimiques marines, y compris des environnements pauvres en nutriments, mais habitables, qui ont favorisé l’adaptation, la compétition et l’évolution des métazoaires au sein du système océanique global (Fig. 3). Alors que les plateaux marins eutrophes accueillaient des démosponges, mais étaient souvent dépourvus du biote d’Ediacara, le bassin salé du sud d’Oman (et les affleurements épais d’Ediacara corrélatifs plus au nord d’Oman) en étant un exemple frappant ; de manière contre-intuitive, les voies maritimes épicontinentales peu profondes, oxiques et moins productives ont été colonisées par le biote d’Ediacara de préférence aux démosponges, malgré la présence de DOM et d’autres détritus organiques disponibles localement pour l’alimentation. On ne sait pas encore si les exigences métaboliques ou la pression sélective environnementale ont restreint le biote multicellulaire à corps mou de l’Édiacarien à ces milieux ; cependant, notre étude souligne que les milieux oligotrophes de l’Édiacarien ont joué un rôle potentiellement crucial dans l’évolution des organismes multicellulaires macroscopiques et l’écologie des communautés marines.