Exceptionally low thermal maturity warstw
Dojrzałość termiczna jest najważniejszym czynnikiem wpływającym na zachowanie starożytnych biomarkerów osadowych, ponieważ cząsteczki organiczne są podatne na zmiany strukturalne i stereochemiczne w trakcie stopniowego zakopywania i przy rosnącym stresie termicznym29. Skały prekambryjskie o odpowiedniej dojrzałości termicznej (dojrzałość okna olejowego lub niższa) są warunkiem koniecznym do zachowania solidnych i pierwotnych zespołów biomarkerów lipidowych. Wielokrotne proporcje stereoizomerów biomarkerów hopanu i steranu, wraz z niezależnymi dowodami w postaci niskich wartości Tmax (głównie w zakresie 417-433 °C, ze średnią 426 °C) z pirolizy Rock-Eval, wskazują, że skały w tym badaniu reprezentują zdecydowanie najbardziej niedojrzałe termicznie skały ediakarskie analizowane do tej pory przy użyciu najnowocześniejszych metod geochemii organicznej (Tabela 1). Mimo że skały te przeszły diagenezę sedymentacyjną, nie weszły znacząco w okno naftowe, z wyjątkiem nieco bardziej dojrzałych próbek 16PL z basenu Podillya w Mołdawii, które nadal nadają się do analizy (dojrzałość od wczesnego do średniego okna naftowego i brak wyraźnych śladów zanieczyszczeń organicznych). Większość zbioru nie była wówczas prawdopodobnie narażona na temperatury grzebania przekraczające 50°C, stąd nasze próbki są wysoce niedojrzałe. Inne linie molekularnych dowodów, które potwierdzają niską dojrzałość termiczną i syngeniczność, obejmują (i) dominację wielopierścieniowych biomarkerów alkanów nad n-alkanami w ekstraktach skalnych (Fig. 2), (ii) przetrwanie wykrywalnych ilości 17β,21β(H)-hopanów, które można oddzielić od bardziej obfitych hopanów posiadających stabilne konfiguracje 17β,21α(H)- i 17α,21β(H)-stereochemiczne (Tab. 1, Supplementary Figs. 1 i 2), (iii) zauważalna nieparzysta-nieparzysta preferencja wśród n-alkanów w zakresie od C22 do C27 (Fig. 2), jako że n-alkany wykazują preferencję co do liczby atomów węgla tylko przed katagenezą38, oraz (iv) generowanie niedojrzałych termicznie biomarkerów hopanu i steranu z (nierozpuszczalnej) fazy kerogenu za pomocą katalitycznej hydrolizy (Supplementary Fig. 2). Niska dojrzałość termiczna osadowej materii organicznej w naszych próbkach jest zgodna z wcześniej opublikowanymi wskaźnikami zmian osadowych, w tym wskaźnikami zmian konodontów i akrytarchów, oraz wcześniejszymi badaniami pirolizy i biomarkerów Rock-Eval dla ediakarskich i paleozoicznych skał osadowych w Baltice36,39,40, jak również mineralogią iłu41. Krytycznie, w środowiskach, gdzie dojrzałość termiczna jest tak niska, mechanizmy znaczącej zmiany pierwotnych sygnałów δ15N i δ13C są w dużej mierze nieobecne42.

Rozkład ekstrahowalnych węglowodorów alifatycznych dla próbki reprezentatywnej. a Chromatogram całkowitych jonów (TIC) dla ekstrahowalnych węglowodorów alifatycznych dla Lugovoe #13-73 m z horyzontu Redkino. Oznaczono serię n-alkanową, prystan (Pr), fitan (Ph) i hopany C27-C34 (oznaczone całkowitą liczbą węgli i stereochemią przy C-17, C-21 i C-22, np. C31αβR); ** oznacza hopeny C29 i C30. Zwraca uwagę dominacja liczebności hopanów nad innymi klasami związków alkanowych. b Częściowy chromatogram jonowy 85-Da wykazuje dominację serii n-alkanów, wykazując wyraźną preferencję nieparzystej liczby atomów węgla w zakresie C22-C27, nad metyloalkanami
Lipid biomarker assemblage patterns
Stosunek obfitości głównych (C27-C35) hopanów do głównych (C27-C29) steranów jest często wykorzystywany do oceny równowagi wkładu bakterii i eukariotycznych organizmów źródłowych do starożytnego ekosystemu wodnego. Hopany są skamieniałościami molekularnymi pochodzącymi od hopanoidów, które są lipidami błon komórkowych syntetyzowanymi przez wiele różnych grup bakterii. Podobnie, sterany pochodzą z prekursorów steroli, które są produkowane prawie wyłącznie przez eukarionty43. Stosunki chmiel/steran (H/St) dla naszych próbek obejmują uderzająco duży zakres wartości od 1,6 do 119,2 (Tabela 1). Dla kontekstu, stosunki H/St z bogatych w substancje organiczne skał neoproterozoicznych i olejów zwykle mieszczą się w wąskim zakresie od 0,5 do 2,030,31,32,33. Podczas gdy istnieje wyraźna różnica w wartościach z młodszego Horyzontu Kotlin (średnia H/St 8,9) i starszego Horyzontu Redkino (średnia H/St 42,9), wartości dla większości próbek są niezwykle wysokie i sugerują anomalnie podwyższony udział bakterii.
Do późnego neoproterozoiku, glony eukariotyczne były ekologicznie znaczącym składnikiem i głównym producentem w wielu ekosystemach morskich15,34,44. Rozbieżność między globalnie ważnym wkładem glonów eukariotycznych w późnym neoproterozoiku, a jednocześnie niskimi poziomami biomarkerów steranu w tych próbkach wskazuje, że musi istnieć jakiś lokalny czynnik determinujący obfitość eukariotów. Najbardziej prawdopodobnym wyjaśnieniem ekstremalnie podwyższonych stosunków hopan/steran obok niskich zawartości całkowitego węgla organicznego (TOC) i niskich wskaźników wodorowych (HI) stwierdzonych w naszych próbkach (Tabela 1) jest to, że warstwy te zostały zdeponowane w oligotroficznych (tj. silnie ograniczonych odżywczo) środowiskach, w których bakterie pokonały glony. Podczas gdy trudno znaleźć współczesne analogi dla starożytnych mórz epikratonicznych powstałych w okresie wysokiego poziomu morza, równoległe obserwacje wyższych stosunków hopan/steran (nawet o rząd wielkości) znaleziono wcześniej dla skał osadowych organicznie czystych w porównaniu z bogatymi w substancje organiczne, osadzonych w ordowicko-siluryjskich epikontynentalnych kanałach morskich45.
Typowo, niska zawartość TOC może być spowodowana ograniczoną depozycją materii organicznej w warunkach niskiej produktywności, niskim potencjałem konserwacyjnym materii organicznej w słupie wody lub osadach, lub rozcieńczeniem przy wysokim strumieniu krzemionkowym. We współczesnych oceanach produktywność jest najczęściej ograniczana przez niski poziom podstawowych składników odżywczych: azotu, fosforu, krzemu i żelaza46. Azot, fosfor i żelazo są ograniczone w regionach otwartego oceanu, gdzie upwelling, wpływ pyłu i spływy przybrzeżne nie dostarczają wystarczających ilości składników odżywczych. We współczesnych środowiskach oligotroficznych stosunek biomasy bakteryjnej do eukariotycznej jest wyższy niż w środowiskach eutroficznych lub mezotroficznych. Co ciekawe, próbki z wychodni Podillya 16PL z horyzontu Redkino w Mołdawii są mułowcami zawierającymi fosforyt i charakteryzują się znacznie niższym stosunkiem hopan/steran (8,1:11,5) niż inne próbki z Redkino w Tabeli 1 (choć nadal znacznie wyższym niż te znalezione w basenie solnym Południowego Omanu lub w bogatych w substancje organiczne osadach fanerozoicznych, które zazwyczaj mieszczą się w wąskim zakresie 0.5-2,0), co może sugerować, że zwiększona dostępność fosforanów mogła korzystnie wpłynąć na stosunek eukariotów do bakterii występujących lokalnie (biorąc pod uwagę, że sygnatury izotopowe azotu są w dużej mierze niezmienne, patrz następna sekcja). Zawartość fosforu (P), jak również stosunki P/Fetotal i P/Al, są ogólnie niskie w późnoediakarskich osadach Podillyi, Ukrainy i Estonii47; z wyjątkiem tego poziomu stratygraficznego, w którym występują konkrecje fosforytowe. Podobnie niskie (~0,01 do ~0,1 mas%) poziomy P w skałach krzemionkowych Kotlin i Redkino odnotowano w rdzeniu wiertniczym z północno-wschodniego marginesu Platformy Wschodnioeuropejskiej11.
Szeroka i płytka topografia w epikontynentalnych morzach Baltica mogła podtrzymywać ograniczenia fosforu lub innych składników odżywczych w warunkach marginalnych, ze względu na autigeniczne wytrącanie fosforanu z minerałami żelaza w utlenionych wodach powierzchniowych i sekwestrację szeregu pierwiastków przez osady szelfowe48. Jeśli fosforan był ograniczającym składnikiem pokarmowym, zwiększenie biodostępnego fosforu zwiększyłoby lokalną produkcję pierwotną i zapewniłoby korzystniejsze warunki wzrostu dla większych rozmiarów jednokomórkowego fitoplanktonu49,50,51. W odniesieniu do współczesnego systemu oceanicznego uznaje się, że morskie pikocyanobakterie, Prochlorococcus i Synechococcus, dominują pod względem liczby komórek fitoplanktonu i biomasy w oligotroficznych tropikalnych i subtropikalnych warunkach oceanicznych, w tym w ograniczonych fosforanami oligotroficznych regionach oceanicznych wód powierzchniowych52. Prochlorococcus i heterotroficzne SAR11 (Pelagibacter) rozkwitają dzięki różnorodnym przystosowaniom, w tym niskim kosztom energii dzięki małym genomom i niskim wskaźnikom replikacji, wyższemu stosunkowi powierzchni do objętości dzięki mniejszym rozmiarom komórek oraz dodatkowym funkcjom wychwytu komórkowego w celu maksymalizacji wykorzystania składników odżywczych53. Wiele bakterii jest również w stanie zastąpić składniki odżywcze o niskiej obfitości, np. poprzez wykorzystanie sulfolipidów zamiast lipidów zawierających fosfor w warunkach niedoboru P54, lub poprzez wykorzystanie alternatywnych substratów, np. pozyskiwanie azotu z atmosferycznego N2 poprzez wiązanie azotu55, w celu złagodzenia stresu związanego ze składnikami odżywczymi. Ostatnie badania dotyczące przetrwania bakterioplanktonu morskiego w środowiskach oligotroficznych56, gdzie dostępny fosforan jest ograniczony, sugerują ważną rolę metabolizmu polifosforanów w morskich oligotrofach. Wybrane eukarionty, które konkurują w środowiskach oligotroficznych, to zazwyczaj małe pikoeukarionty, które mogą uzupełniać swoje potrzeby żywieniowe poprzez miksotrofię57. Stosunek biomasy picoeukariotów do biomasy cyjanobakterii ma tendencję do zwiększania się w warunkach zwiększonej podaży składników odżywczych49. Możliwy wpływ fosforu i innych biolimitujących składników pokarmowych w moderowaniu produktywności pierwotnej i struktury zbiorowisk morskich w epeirycznych zatokach ediakarskich wymaga dalszych badań.
Bez wyjątku, obfitość steranów C29 jest większa niż odpowiadających im steranów C27 lub C28 we wszystkich naszych lokalizacjach (Tabela 1). Przewaga C29 nad C27 i C28 steranami prawdopodobnie wskazuje na dominację zielenic w społeczności fitoplanktonu eukariotycznego44,58. Cecha ta została zaobserwowana w większości wcześniejszych badań biomarkerów ediakarskich15,30,31,32,33,34,35,36,44. Warto zauważyć, że rozkład steranu C30 w kilku próbkach z każdego rdzenia wiertniczego w Kotlin Horizon zawiera niskie, ale wykrywalne ilości biomarkera steranu z demospondy, znanego jako 24-izopropylocholestan (24-ipc)59,60. W sumie sterany 24-ipc odnotowano w próbkach skał i ropy naftowej pochodzących aż z kriogenu (>635 mln lat temu) w basenie solnym południowego Omanu i stanowią one najstarsze dowody biomarkerów lipidowych dla metazoanów59,60. Biomarkery 24-ipc w naszych próbkach były albo około jeden rząd wielkości niższe w obfitości (w stosunku do całkowitych stosunków C27 do C30 steranu, były one tylko 0,06-0,61%; średnia = 0,22%) w porównaniu z ediakarskimi skałami i olejami z Południowego Omanu (średnio 1,7%59) lub były poniżej granic wykrywalności ze względu na pomijalną obfitość dla większości próbek.
Stosunki izotopów azotu i węgla organicznego
Izotopy azotu mogą pomóc w rozeznaniu względnej równowagi w cyklu azotowym oraz stopnia, w jakim wiązanie azotu lub niekompletna denitryfikacja były znaczącymi ścieżkami wpływającymi na równowagę składników odżywczych dostępnych dla społeczności morskich. Gdy bakterie diazotroficzne wiążą azot cząsteczkowy z powodu braku azotu związanego w słupie wody, może to prowadzić do uzyskania wartości izotopów azotu w osadzie bliskich 0‰61. Pozostaje to w sprzeczności z dodatnimi znakami izotopów azotu (w zakresie od +2 do +10‰) z trybem od +4 do +6‰ stwierdzonymi dla neoproterozoicznych osadów morskich zdeponowanych w warunkach, które uważa się za warunki ubogie w azotany, w których azotan uległ jedynie częściowej denitryfikacji62. Wartości izotopów azotu dla wszystkich naszych próbek z wyjątkiem jednej odstającej (Tabela 1) obejmują ograniczony dodatni zakres od +3,5 do +6,5‰, który pokrywa się z trybem dla późno neoproterozoicznych skał osadowych bogatych w materię organiczną62. Ograniczone przez proxy redoks sugerujące, że panowały tlenowe warunki w kolumnie wodnej, nasze dane sugerują, że azotan dominował w puli rozpuszczonego azotu nieorganicznego. Zakres wartości δ15N sugeruje również, że wiązanie N2 nie było głównym sposobem pozyskiwania azotu przez producentów pierwotnych. Cykl azotowy był raczej zdominowany przez recykling w kolumnie wodnej, a organiczny N podlegał ilościowemu utlenianiu do azotanu, tak jak ma to miejsce we współczesnych, proksymalnych środowiskach morskich. Denitryfikacja bentosowa i pochówek organicznego N byłyby głównymi pochłaniaczami rozpuszczonego azotu nieorganicznego, a zakres wartości δ15N sugeruje jedynie ograniczoną rolę niekompletnej denitryfikacji w słupie wody, która zwykle powoduje znaczne wzbogacenie 15N63. Dlatego ograniczona zmienność w wartościach δ15N prawdopodobnie odzwierciedla δ15N azotanów przenoszonych na platformę, z niewielkim wpływem ograniczonej redukcji azotanów w słupie wody i wiązania N2. Bez silnych dowodów δ15N dla wiązania azotu, hipotezujemy, że azot nie był głównym biolimitującym składnikiem odżywczym w epicontinentalnych basenach Baltica. Izolacja od rzecznych i eolicznych źródeł fosforu oraz warunki tlenowe w szerokich, płytko-morskich basenach epikontynentalnych mogły wzmocnić usuwanie autigenicznego fosforytu i faz zawierających metale śladowe, powodując ograniczenie składników pokarmowych, które ograniczały wzrost i produkcję komórek eukariotycznych.
Całkowite wartości izotopów węgla organicznego (CTOC) wahają się od -23,0 do -33,9‰, z największymi różnicami obserwowanymi pomiędzy różnymi lokalizacjami rdzeni wiertniczych. Względne wzbogacenie 13C do stosunków izotopowych wyższych niż około -28‰ kontrastuje z danymi z równoległych warstw osadzonych w eutroficznych, otwartych środowiskach morskich Omanu30,31. Jednakże zakres, który zgłaszamy jest ogólnie zgodny z zakresem δ13CTOC dla innych lokalizacji z Baltica osadzonych w tym samym okresie czasu11. Różnica między Baltica i Oman może podkreślić fakt, że wartości δ13CTOC nie wyłącznie odzwierciedlają jednolite zmiany sekularne w cyklu węglowym w tym przedziale czasu31,64. Raczej, zakres δ13CTOC z Baltica może, w części, odzwierciedlać bakteryjnie zdominowaną ekologię mikrobiologiczną sugerowaną przez unikalne stosunki biomarkerów lipidowych. Małe rozmiary komórek, wysoki stosunek powierzchni do objętości i powolne tempo wzrostu w warunkach oligotroficznych mogą zwiększyć wielkość frakcjonowania podczas autotrofii (εp)65,66, skutkując niskimi wartościami δ13C.
Mechanizm dla bardziej wzbogaconych w 13C sygnatur izotopowych w zakresie podanym11 dla późnoediakarskiej sukcesji Baltica jest mniej jasny. Potencjalny wkład detrytycznej, metamorficznie zmienionej materii organicznej do prekambryjskich sukcesji osadowych o niskim TOC jest możliwym mechanizmem dla różnic pomiędzy miejscami w δ13CTOC64. Jednakże zawartość TOC w naszych próbkach, choć niska, jest generalnie większa niż 0.10% mas, nie ujawniając zależności między zawartością TOC a wartościami δ13C. Ponadto, nasze próbki zawierają niedojrzałą termicznie materię organiczną i wzory lipidów biomarkerów, które nie są zgodne z głównie allochtonicznym źródłem węgla. Bardziej wzbogacone w 13C sygnatury δ13CTOC mogą być wynikiem alternatywnych mechanizmów asymilacji węgla. Mechanizmy koncentracji węgla lub aktywnego wychwytu wodorowęglanów przez prokariota mogą skutkować mniejszymi wartościami εp i wyższymi wartościami δ13CTOC67,68. Pojawienie się ediakarskiej fauny i flory mogło znacznie rozszerzyć morskie sieci pokarmowe i pobudzić nowe drogi heterotrofii mikrobiologicznej, w tym możliwy wkład złożonego obiegu węgla w matach mikrobiologicznych na dnie. Z dodatkowym uwzględnieniem potencjalnie ważnej roli rozpuszczonej materii organicznej69 jako źródła węgla, szeroki zakres wartości δ13CTOC prawdopodobnie odzwierciedla szereg biogeochemicznych procesów obiegu węgla, które mogą być unikalne dla ewoluującego środowiska morskiego Ediacaran i mogą być związane ze zjawiskiem, dla którego nie mamy reprezentatywnych współczesnych analogów.
Wczesne zespoły biomarkerów lipidowych i wskaźniki stabilnych izotopów dla bentosowej, mikrobiologicznej produkcji mat w tandemie ze stałym ograniczeniem azotu zostały znalezione we wczesnych skałach triasowych z południowych Chin70. W następstwie masowego wymierania pod koniec permu w Meishan nastąpił duży skok w stosunkach hopan/steran (do ok. 60), któremu towarzyszyło silne przesunięcie sygnatury izotopowej Norg do wartości od 0 do -2‰, spójne z diazotrofią bakteryjną. Wiąże się to z wysokim sygnałem 2-metylopropanów (indeks 2-metylopropanowy do 33%) i charakterystycznych metyloalkanów, które wskazują na proliferację mat mikrobiologicznych na dnie. Nie obserwujemy podobnego trendu w sygnaturze izotopu azotu lub wzorców biomarkerów, które w oczywisty sposób wskazywałyby na znaczący wkład maty mikrobiologicznej w naszym ediakarskim zestawie danych, chociaż w naszych próbkach znaleziono pojedynczą wartość zubożoną w 15N (Tabela 1). Dlatego silny sygnał bakteryjny obserwowany w naszych próbkach Baltica prawdopodobnie nie jest w przeważającej mierze sygnaturą bentosowych mat mikrobiologicznych.
Paleośrodowiskowe utrzymanie fauny i flory Ediacara vs. demosponges
. demosponges
Widoczne oligotroficzne warunki w epikratonicznych i kontynentalnych basenach Baltica, gdy dryfowała od wysokich do niskich szerokości geograficznych od późnego ediakaru do wczesnego kambru, mogą być związane albo z ograniczoną adwekcją względnie bogatych w składniki odżywcze wód głębokich, albo zubożeniem w składniki odżywcze, wynikającym z asymilacji i wymiatania podczas transportu i depozycji w tych szerokich, płytko-morskich basenach epikontynentalnych (Fig. 3). Baseny te były epizodycznie izolowane od oceanów i rozwijały się w nich warunki hipersolalności (np. w okresie Redkino) i słonawości (np. w okresie Kotlin71). Długotrwała stabilność tektoniczna zaowocowała niską topografią późnoediakarskiej Baltiki, bardzo podatnej na zalewanie i nieefektywne dostarczanie fosforu pochodzącego z wietrzenia. W przeciwieństwie do Baltica, badania biomarkerów warstw z supergrupy Huqf w Omanie wykazały istnienie eutroficznego ekosystemu, bogatego w mikroalgi30,31,59, ale pozbawionego fauny i flory Ediacara, nawet w rozległych wychodniach w Górach Omańskich i regionie Huqf, w położeniu od wewnętrznego do zewnętrznego szelfu3. Paleogeografia Basenu Solnego Południowego Omanu została zrekonstruowana dla późnego neoproterozoiku na ~13° od równika na półkuli południowej, zasadniczo podobna do paleolatitudy Baltica72.
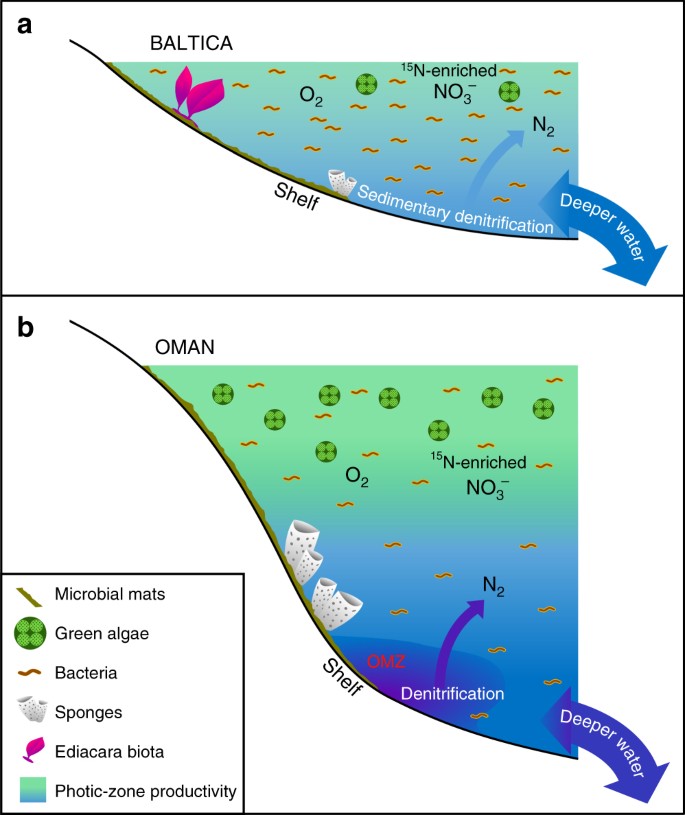
Główne różnice w mało produktywnych vs. produktywnych ediakarskich środowiskach morskich. Schematy pokazane są dla a rozległych oligotroficznych i płytkich epikontynentalnych basenów Baltica często zdominowanych przez bakteryjną produktywność, gdzie fauna Ediacara o miękkich ciałach rozkwitała i gdzie denitryfikacja i anammox prawdopodobnie były ograniczone do osadów; oraz b eutroficzne i głębsze morskie środowisko szelfowe w basenie solnym Południowego Omanu, gdzie zielenice kwitły jako producenci pierwotni i obficie występowały gąbki, ale fauna miękkociernista Ediacara nie była widoczna (skamieniałości fauny Ediacara nie występują również w korelujących z nimi ediakarskich wychodniach w północnym Omanie). Na produktywnych obrzeżach kontynentalnych denitryfikacja i anammox prawdopodobnie zachodziły zarówno w słupie wody, jak i w osadach
Nie ma dowodów na trwałą i rozległą anoksję na płytkich obrzeżach kontynentalnych Baltiki i wokół nich podczas późnego Ediakaru11, i sugerujemy, że warunki oligotroficzne opisane tutaj zostały spowodowane przez nieefektywne lądowych i głębinowych strumieni składników odżywczych do tych ustawień, zasadniczo podobne do tych w oligotroficznych środowiskach we współczesnych systemach oceanicznych. Istotnie, badania geochemiczne pierwiastków śladowych z Utkina Zavod i przyległych rdzeni wiertniczych z obszaru St. Petersburga sugerują, że nasze próbki zostały zdeponowane w warunkach tlenowych73. Interpretacja ta jest niezależnie wspierana przez nasze dane z rozszerzonego rozkładu hopanów (C31-C35), których obfitość gwałtownie spada powyżej związków C31 wraz ze wzrostem liczby atomów węgla, co jest charakterystyczne dla degradacji łańcucha bocznego bakteriohopanepolioli podczas diagenezy w warunkach tlenowych29, a także przez niskie wskaźniki wodorowe, mierzone metodą pirolizy Rock-Eval (wszystkie są niższe niż 230 mg/g TOC, a wiele z nich jest niższych niż 100 mg/g TOC) dla tych niedojrzałych próbek, co wynika z tworzenia się rekalcytującego kerogenu poprzez degradację oksydacyjną i rekondensację ubogiej w lipidy biomasy pierwotnej w środowiskach lokalnie tlenowych (Tabela 1). Wielokomórkowe organizmy ediakarskie, które istniały w tych płytkich środowiskach morskich, musiały mieć wystarczającą ilość substratów organicznych dla heterotrofii, by zaspokoić swoje potrzeby pokarmowe, a także wystarczającą ilość tlenu i innych składników odżywczych, by podtrzymać swój metabolizm. Epikontynentalne baseny Baltica były prawdopodobnie bardziej trwale utlenione niż wysoce produktywne środowiska, które otaczały strefy minimum tlenowego, takie jak na środkowym i zewnętrznym szelfie Południowego Basenu Solnego Omanu, gdzie oddychanie obfitej biomasy planktonowej utrzymywałoby niższy poziom rozpuszczonego tlenu poniżej strefy fotycznej.
Ediacara biota i inne wielokomórkowe organizmy żyjące u wybrzeży Baltica musiałyby radzić sobie ze zmieniającymi się źródłami pożywienia (bakteryjne vs eukariotyczne), ponieważ strumienie składników odżywczych zmieniały się w czasie, w tym małe komórki i detrytus organiczny w basenach epikontynentalnych, gdzie bakterie były dominującymi producentami pierwotnymi (Fig. 3). Współczesne zbiorowiska raf koralowych przetrwały w tropikalnym środowisku oligotroficznym dzięki skutecznemu recyklingowi składników pokarmowych, w tym dzięki generowaniu strumienia rozpuszczonej materii organicznej (DOM), który pomaga utrzymać heterotrofię fauny w ekosystemie rafy74. Ustanowienie morskiej struktury troficznej z wielokomórkowymi organizmami eukariotycznymi utrzymującymi się dzięki odżywianiu detrytusem organicznym musiało poprzedzać ekspansję środowiskową eukariotów na skalę globalną do zróżnicowanych środowisk morskich, która nastąpiła w interwale tonian-kryogen (ok. 800-635 mln lat temu), jak wynika z zapisów biomarkerów15. Moglibyśmy zatem oczekiwać, że dostrzeżemy dowody na znaczące różnice w morskiej społeczności i strukturze troficznej w różnych miejscach w późnym okresie ediakaru, gdzie lokalna równowaga składników odżywczych wybierała społeczności drobnoustrojów bogate w eukariotę lub pozbawione eukariotów, a stopniowa ekspansja organizmów wielokomórkowych dodawała kolejny wymiar złożoności na poziomie organizmów i społeczności. Znaczny regionalny kontrast w dostępności fosforanów i innych składników odżywczych w środowiskach szelfowych jest również oczekiwaną konsekwencją niejednorodnej globalnej morskiej struktury redoks w późnym Ediakarze, przed natlenieniem głębokiego oceanu38.
Dominacja bakteryjnej produktywności pikoplanktonu i związanych z nią produktów degradacji rozpuszczonej materii organicznej (DOM) w epikontynentalnych basenach Baltica mogła podtrzymywać ekosystem pętli mikrobiologicznej równolegle z konwencjonalną strukturą troficzną opartą na większych producentach planktonowych50. Mogło to sprzyjać różnym sposobom heterotrofii, w tym żywieniu w zawiesinie i, być może, osmotrofii, jako realnej strategii żywieniowej dla niektórych rangeomorfów, gąbek i innych wielokomórkowych organizmów późnego ediakaru18 w połączeniu z pojawiającą się aktywną (ruchliwą) heterotrofią19,20. Systemy wodne o ograniczonej zawartości składników odżywczych są z reguły często zdominowane przez mały jednokomórkowy fitoplankton i plankton heterotroficzny, z bakteriami prześcigającymi eukarionty i utrzymującymi niską biomasę netto w oligotroficznych środowiskach morskich współczesnego oceanu49,51. DOM jest ważnym źródłem organicznych składników odżywczych i często kontroluje produktywność i biomasę netto we współczesnych oligotroficznych morzach tropikalnych, ale podobnie jak współlimitacja azotu i fosforu75. Podobnie, DOM był prawdopodobnie również ważnym substratem dla podtrzymania heterotroficznych bakterii i pętli mikrobiologicznej w starożytnych środowiskach oligotroficznych. Bentosowe maty mikrobiologiczne mogły być składnikiem tej zdominowanej przez bakterie sieci pokarmowej, a podstawy mat zostały wskazane jako czynnik ułatwiający zachowanie fauny i flory Ediacara w środowisku morskim7. Podczas gdy umiarkowane wartości indeksu 2-metylopropanowego zostały znalezione dla podzbioru naszych próbek (4-10%, Tabela 1), bardzo niska obfitość metyloalkanów w stosunku do n-alkanów (Rys. 1) oraz tylko śladowe i sporadyczne występowanie karotenoidów sugeruje, że maty mikrobiologiczne nie zdominowały pierwotnej produktywności76 , a bakterie planktonowe i, odpowiednio, ich produkty rozpadu DOM kwitły. Ogólnie niskie wartości indeksu wodorowego (Tabela 1; szczególnie dla próbek z Redkino), pomimo niskiej dojrzałości termicznej warstw i zespołów biomarkerów, są również spójne z ubogim w lipidy wkładem organicznym w głównie tlenowych, płytko-morskich środowiskach marginalnych. Występowanie maty mikrobiologicznej, oceniane jedynie na podstawie tekstury osadów, nie mówi nam o równowadze między eukariotami (mikroalgami) a bakteriami w każdym przypadku, ani też nie jest w stanie określić względnego udziału planktonu mikrobiologicznego w ogólnej produktywności pierwotnej i utrzymaniu sieci pokarmowych. Zbiorowiska maty mikrobiologicznej, które często zawierają obfite eukarionty, a węglany z Grupy Ara z południowego Omanu z wyraźnymi trombolitycznymi i kruchymi laminami są dobrym przykładem późnych środowisk neoproterozoicznych ze znaczącym udziałem maty mikrobiologicznej, dającym obfite sygnały steranowe z powodu dużego udziału mikroalg30,59.
Niezwykle niski poziom biomarkerów 24-ipc sugeruje, że choć demospongi były czasem obecne, to w tych środowiskach były nieliczne. Może to wskazywać na mniejsze możliwości dla mniejszych zwierząt filtrujących, konkurujących z fauną i florą Ediacara w tych mało produktywnych środowiskach, niewystarczającą odporność demospongów na bardziej energetyczne warunki płytkowodne powyżej podstawy falowania w czasie pogody lub lepsze możliwości adaptacyjne gąbek do warunków niskotlenowych dynamicznie utrzymywanych poniżej strefy fotycznej w środowiskach eutroficznych (Rys. 3). W sumie, 24-ipc nie został wykryty w większości próbek z horyzontu Redkino, które również mają generalnie najwyższe współczynniki H/St wśród naszego zestawu próbek, co być może wskazuje na ekologiczną zmianę w czasie depozycji horyzontu Kotlin. Alternatywnie, jest również prawdopodobne, że gąbki zamieszkiwały te środowiska, ale nie produkowały obficie tych diagnostycznych biomarkerów steroidowych, choć wydaje się to mniej prawdopodobne, gdyż 24-ipc steran należy do najczęściej wykrywanych związków C30 steranu w warstwach ediakarskich i olejach30,3133,59,60 i jest również wykrywalny w podzbiorze naszych próbek (Tabela 1).
Pomimo, że trwałe oligotroficzne środowiska morskie sugerowane przez nasze dane reprezentują lokalne warunki w ediakarskich oceanach, prawdopodobnie nie były one rzadkością w prekambryjskich płytkich morskich kanałach (Fig. 3). Zdecydowana większość wcześniejszych badań biomarkerów ediakarskich została przeprowadzona na bogatych w substancje organiczne skałach osadowych osadzonych w środowiskach eutroficznych i ich produktach ropopochodnych, które generalnie dają biomarkery zgodne ze znaczącym udziałem mikroglonów44. Nasze wyniki podkreślają znaczenie badania szerszej gamy środowisk depozycyjnych, w tym warstw organiczno-materiałowych o odpowiedniej dojrzałości termicznej i różnych litologiach15,3034,59, w celu uzyskania dokładniejszego obrazu skali heterogeniczności chemii i ekologii morskiej z miejsca na miejsce. Pomimo postępującego wietrzenia oceanu i zwiększonego wietrzenia chemicznego oraz dostarczania składników odżywczych podczas rozpadu Rodinii i przez cały okres ediakarski77,78, heterogeniczność oceanu utrzymywała różnorodność morskich warunków chemicznych, w tym ubogie w składniki odżywcze, ale nadające się do zamieszkania środowiska, które sprzyjały adaptacji, konkurencji i ewolucji metazoanów w globalnym systemie oceanicznym (Fig. 3). Podczas gdy eutroficzne szelfy morskie gościły demospongi, ale często brakowało na nich fauny Ediacara, czego dobitnym przykładem jest South Oman Salt Basin (i korelujące z nim grube wychodnie ediakaru dalej na północ w Omanie); co ciekawe, płytkie, tlenowe i mniej produktywne epikontynentalne szlaki morskie były kolonizowane przez faunę Ediacara w przeciwieństwie do demospongów, mimo że DOM i inny detrytus organiczny był lokalnie dostępny do żerowania. Nie wiadomo, czy wymagania metaboliczne lub presja selekcyjna środowiska ograniczyły wielokomórkową faunę Ediacaran o miękkich ciałach do tych środowisk, jednak nasze badania podkreślają, że ediakarskie środowiska oligotroficzne odegrały potencjalnie kluczową rolę w ewolucji makroskopowych organizmów wielokomórkowych i ekologii społeczności morskich.
.