Exceptionellt låg termisk mognad i strata
Termisk mognad är den enskilt viktigaste faktorn som påverkar bevarandet av gamla sedimentära biomarkörer eftersom organiska molekyler är känsliga för strukturella och stereokemiska förändringar under progressiv nedgrävning och med ökande termisk stress29. Prekambriska bergarter med lämplig termisk mognad (oljefönstermognad eller lägre) är en förutsättning för att bevara robusta och primära lipidsammansättningar av biomarkörer. Flera stereoisomerförhållanden för biomarkörer av hopan och steran, tillsammans med oberoende bevis från låga Tmax-värden (mestadels inom intervallet 417-433 °C, med ett medelvärde på 426 °C) från Rock-Eval-pyrolys, indikerar att bergarterna i den här studien representerar de överlägset mest termiskt omogna ediacariska bergarterna som hittills har analyserats med hjälp av de senaste metoderna för organisk-geokemisk analys (tabell 1). Även om våra stenar har genomgått sedimentär diagenes har de inte passerat signifikant in i oljefönstret, bortsett från de något mer mogna 16PL-proverna från Podillya-bassängen i Moldavien, som fortfarande lämpar sig för analys (tidig till medellång mognad i oljefönstret och inga uppenbara tecken på organiska föroreningar). Majoriteten av uppsättningen hade då troligen inte utsatts för begravningstemperaturer på över 50 °C, varför våra prover är mycket omogna. Andra linjer av molekylära bevis, som stöder låg termisk mognad och syngenicitet, inkluderar (i) en dominans av polycykliska biomarkörer alkaner över n-alkaner i bergsextrakt (fig. 2), (ii) överlevnad av detekterbara mängder 17β,21β(H)-hopaner som kan upplösas från de mer abundanta hopanerna som besitter stabila 17β,21α(H)- och 17α,21β(H)-stereokemiska konfigurationer (tabell 1, supplementära figurer. 1 och 2), iii) en märkbar odd-over-even-preferens bland n-alkanerna i intervallet C22-C27 (fig. 2), eftersom n-alkaner visar en kolnummerpreferens endast före katagenesen38 , och iv) generering av termiskt omogna biomarkörer av hopaner och steraner från den (olösliga) kerogenfasen med hjälp av katalytisk hydropyrolys (kompletterande fig. 2). Den låga termiska mognaden hos sedimentärt organiskt material i våra prover stämmer överens med tidigare publicerade indikatorer på sedimentär alteration, inklusive konodont- och akritarksalterationsindex, och tidigare Rock-Eval-pyrolys- och biomarkörstudier av Ediacaran och paleozoiska sedimentära bergarter i Baltica36,39,40, samt lermineralogi41. Kritiskt är att i miljöer där den termiska mognaden är så här låg är mekanismerna för betydande förändring av primära δ15N- och δ13C-signaler i stort sett frånvarande42.

Fördelning av extraherbara alifatiska kolväten för ett representativt prov. a Totaljonkromatogram (TIC) för extraherbara alifatiska kolväten för Lugovoe #13-73 m från Redkino Horizon. N-alkanserien, pristan (Pr), fytan (Ph) och C27-C34-hopaner (betecknade med deras totala kolantal och stereokemi vid C-17, C-21 och C-22, t.ex. C31αβR) är märkta; ** betecknar C29- och C30-hopaner. Observera att hopaner dominerar över andra klasser av alkanföreningar. b Partiellt 85-Da jonkromatogram visar dominans av n-alkanserien, som uppvisar en tydlig preferens för udda och jämna kolnummer i C22-C27-området, över metylalkaner
Mönster för sammansättning av biomarkörer för lipider
En abundansförhållande mellan de viktigaste (C27-C35) hopanerna och de viktigaste (C27-C29) steranerna används ofta för att bedöma balansen mellan bakteriell och eukaryotisk tillförsel av källorganismer till det forntida akvatiska ekosystemet. Hopaner är molekylära fossiler som härrör från hopanoider, som är cellmembranlipider som syntetiseras av en mängd olika bakteriegrupper. På samma sätt härstammar steraner från sterolprekursorer, som nästan uteslutande produceras av eukaryoter43. Förhållandet mellan hopaner och steraner (H/St) för våra prover täcker ett slående stort intervall av värden från 1,6 till 119,2 (tabell 1). I ett sammanhang kan nämnas att H/St-förhållanden från organiska rika neoproterozoiska bergarter och oljor vanligtvis faller inom ett smalt intervall från 0,5 till 2,030,31,32,33. Även om det finns en tydlig skillnad mellan värdena från den yngre Kotlinhorisonten (genomsnittlig H/St på 8,9) och den äldre Redkinohorisonten (genomsnittlig H/St på 42,9) är värdena för de flesta proverna ovanligt höga och tyder på onormalt höga bidrag från bakterier.
I slutet av neoproterozoikum var eukaryotiska alger en ekologiskt betydelsefull komponent och stora producenter i många marina ekosystem15,34,44. Diskrepansen mellan det globalt viktiga bidraget från eukaryota alger i slutet av neoproterozoikum och ändå de låga nivåerna av steranbiomarkörer i dessa prover tyder på att det måste finnas någon lokal bestämningsfaktor för eukaryota abundans. Den mest förenklade förklaringen till de extremt förhöjda förhållandena mellan hopan och steran tillsammans med låga halter av totalt organiskt kol (TOC) och låga väteindex (HI) som vi fann i våra prover (tabell 1) är att dessa lager avlagrades i oligotrofa (dvs. starkt näringsbegränsade) miljöer, där bakterier utkonkurrerade algerna. Även om det är svårt att hitta moderna analoger för gamla epikratoniska hav som utvecklades under hög havsnivåställning, har man tidigare funnit parallella observationer av högre hopan/steranförhållanden (med upp till en storleksordning) för organiskt magra jämfört med organiskt rika sedimentära bergarter som avlagrats i epikontinentala havsområden från Ordovicium-Silurien45.
Typiskt kan låg TOC-halt orsakas av begränsad deponering av organiskt material i miljöer med låg produktivitet, låg konserveringspotential hos organiskt material i vattenkolumnen eller sedimenten, eller utspädning med ett högt siliklastiskt flöde. I moderna hav begränsas produktiviteten oftast av låga nivåer av de essentiella näringsämnena kväve, fosfor, kisel och järn46. Kväve, fosfor och järn begränsas i öppna havsområden där uppvandring, stofttillförsel och avrinning från kusten inte tillför tillräckliga mängder näringsämnen. I moderna oligotrofa miljöer är förhållandet mellan bakteriell och eukaryotisk biomassa högre än i eutrofa eller mesotrofa miljöer. Intressant nog är Podillya 16PL-utfallsproverna från Redkinohorisonten i Moldavien fosforithaltiga lerstenar och ger betydligt lägre hopan/steran-förhållanden (8,1:11,5) än de andra Redkino-proverna i tabell 1 (även om de fortfarande är betydligt högre än de som hittats i Saltbassängen i södra Oman eller i organiska rika fanerozoiska sediment, som vanligen ligger inom ett snävt intervall på 0,1:11,5).5-2,0 intervall), vilket möjligen tyder på att ökad fosfattillgänglighet kan ha påverkat förhållandet mellan eukaryoter och bakterier som hittats lokalt på ett gynnsamt sätt (med tanke på att kväveisotopsignaturerna i stort sett är invarianta, se nästa avsnitt). Fosforinnehållet (P), liksom förhållandet P/Fetotal och P/Al, är annars generellt sett lågt för de sena ediakariska sedimenten i Podillya, Ukraina och Estland47; med undantag för denna stratigrafiska nivå som är markerad med fosforitnoduler. På samma sätt rapporterades låga (~0,01 till ~0,1 viktprocent) halter av P i siliklastiska bergarter från Kotlin och Redkino från en borrkärna från den nordöstra marginalen av den östeuropeiska plattformen11.
Den breda och grunda topografin i de epikontinentala haven över Baltica kan ha upprätthållit begränsningar av fosfor eller andra näringsämnen i marginella miljöer, på grund av authigen utfällning av fosfat med järnmineraler i det oxiska ytvattnet och sekvensering av en rad grundämnen av shelfsediment48. Om fosfat var ett begränsande näringsämne skulle en ökning av biotillgänglig fosfor ha ökat den lokala primärproduktionen och gett gynnsammare tillväxtförhållanden för större encelliga fytoplankton49,50,51. När det gäller det moderna havssystemet är det känt att de marina picocyanobakterierna Prochlorococcus och Synechococcus dominerar antalet celler och biomassa hos fytoplankton i oligotrofa tropiska och subtropiska havsmiljöer, inklusive fosfatbegränsade oligotrofa områden i havets ytvatten52. Prochlorococcus och den heterotrofa SAR11 (Pelagibacter) blomstrar tack vare en rad olika anpassningar, bland annat låga energikostnader på grund av små genomer och låg replikationshastighet, ett högre förhållande mellan yta och volym genom mindre cellstorlekar och ytterligare cellupptagningsfunktioner för att maximera utnyttjandet av näringsämnen53. Många bakterier kan också ersätta näringsämnen som finns i liten mängd, t.ex. genom att använda sulfolipider i stället för fosforbärande lipider i P-bristmiljöer54 , eller genom att använda alternativa substrat, t.ex. genom att hämta kväve från atmosfäriskt N2 via kvävefixering55 , för att lindra näringsämnesstress. En nyligen genomförd studie om överlevnad av marint bakterioplankton i oligotrofa miljöer56 , där den tillgängliga fosfatmängden är begränsad, tyder på en viktig roll för polyfosfatmetabolismen i marina oligotrofa miljöer. De utvalda eukaryoter som konkurrerar i oligotrofa miljöer är vanligtvis små picoeukaryoter, som kan komplettera sina näringsbehov genom mixotrofi57. Förhållandet mellan picoeukaryoter och cyanobakteriernas biomassa tenderar att öka vid ökad tillförsel av näringsämnen49. Det möjliga inflytandet av fosfor, och andra biolimiterande näringsämnen, i modereringen av primärproduktiviteten och den marina samhällsstrukturen i epeiriska havsområden från Ediacaran kräver ytterligare undersökningar.
Och utan undantag är abundansen av C29-steraner större än motsvarande C27- eller C28-steraner för alla våra platser (tabell 1). En övervikt av C29-steraner jämfört med C27- och C28-steraner indikerar sannolikt en grönalgdominans inom det eukaryota fytoplanktonsamhället44,58. Detta särdrag har observerats i de flesta tidigare studier av biomarkörer från Ediacaran15,30,31,32,33,34,35,36,44. Särskilt noterbart är att C30-steranfördelningen i flera prover från varje borrkärna i Kotlin Horizon innehåller låga, men påvisbara, mängder av demosponge-steranbiomarkören känd som 24-isopropylcholestan (24-ipc)59,60. Totalt sett har 24-ipc-steraner rapporterats i sten- och oljeprover från så långt tillbaka som kryogenian (>635 Ma sedan) i Saltbassängen i södra Oman och representerar de äldsta bevisen på lipidbiomarkörer för metazoer59,60. 24-ipc-biomarkörerna i våra prover var antingen omkring en storleksordning lägre i abundans (i förhållande till de totala C27- till C30-steranförhållandena var dessa endast 0,06-0,61 %; medelvärde = 0,22 %) jämfört med ediacariska stenar och oljor från södra Oman (i genomsnitt 1,7 %59) eller låg under detektionsgränserna på grund av försumbar abundans för majoriteten av proverna.
Väve- och organiskt kolisotopförhållanden
Väveisotoper kan hjälpa till att urskilja den relativa balansen i kvävecykeln och i vilken grad antingen kvävefixering eller ofullständig denitrifikation var betydande vägar för att påverka den näringsbalans som är tillgänglig för marina samhällen. När diazotrofa bakterier fixerar molekylärt kväve på grund av brist på fixerat kväve i vattenpelaren kan detta ge sedimentära bulkkväveisotopvärden nära 0‰61. Detta står i kontrast till de positiva kväveisotopsignaturer (i intervallet +2 till +10‰) med ett läge på +4 till +6‰ som hittats för de neoproterozoiska marina sediment som avsatts under vad som anses vara nitratrika förhållanden där nitratet endast har genomgått en partiell denitrifikation62. Kväveisotopvärdena för alla våra prover med undantag för ett utfall (tabell 1) täcker ett begränsat positivt intervall från +3,5 till +6,5‰, vilket överlappar med modus för sena neoproterozoiska sedimentära bergarter som är rika på organiskt material62. Begränsat av redoxproxier som tyder på att det rådde oxiska förhållanden i vattenpelaren, innebär våra data att nitrat dominerade den upplösta oorganiska kvävepoolen. Intervallet av δ15N-värden tyder också på att N2-fixering inte var den primära metoden för kväveanskaffning för primärproducenter. Kvävecykeln dominerades sannolikt snarare av återvinning i vattenkolumnen, och organiskt kväve var föremål för kvantitativ oxidation till nitrat, vilket det är i moderna, proximala marina miljöer. Benthisk denitrifikation och nedgrävning av organiskt kväve skulle ha varit de primära sänkorna för löst oorganiskt kväve, och intervallet av δ15N-värden tyder på att den ofullständiga denitrifikationen i vattenpelaren, som vanligtvis resulterar i en betydande 15N-anrikning, endast spelade en begränsad roll63. Den begränsade variationen i δ15N-värdena återspeglar därför troligen δ15N för nitrat som advekteras till plattformen, med mindre påverkan från begränsad nitratreduktion och N2-fixering i vattenkolumnen. Utan starka δ15N-bevis för kvävefixering antar vi att kväve inte var det primära biolimiterande näringsämnet i de epikontinentala bassängerna i Baltica. Isolering från flod- och eoliska fosforkällor och oxiska förhållanden i de breda, grunda marina epikontinentala bassängerna kan ha förbättrat avlägsnandet av authigen fosforit och spårmetallförande faser, vilket resulterade i en näringsbegränsning som begränsade eukaryotisk celltillväxt och produktion.
Värden av isotoper för totalt organiskt kol (CTOC) varierar mellan -23,0 och -33,9 ‰, och de största skillnaderna observerades mellan de olika borrkärnepositionerna. Den relativa 13C-anrikningen till isotopförhållanden högre än ca -28‰ står i kontrast till data från samtida lager som avlagrats i eutrofa, öppna marina miljöer i Oman30,31. Det intervall som vi rapporterar är dock generellt sett förenligt med δ13CTOC-området för andra platser från Baltica som avlagrats under samma tidsperiod11. Skillnaden mellan Baltica och Oman kan belysa det faktum att δ13CTOC-värdena inte uteslutande återspeglar en enhetlig sekulär förändring i kolcykeln under detta tidsintervall31,64. Snarare kan δ13CTOC-intervallet från Baltica delvis återspegla den bakteriellt dominerade mikrobiella ekologin som antyds av de unika förhållandena för lipidbiomarkörer. Små cellstorlekar, höga förhållanden mellan yta och volym och långsam tillväxt under oligotrofa förhållanden kan öka storleken på fraktioneringen under autotrofi (εp)65,66, vilket resulterar i låga δ13C-värden.
Mekanismen för de mer 13C-berikade isotopiska signaturerna inom intervallet som rapporterats11 för Baltica-successionen från sent Ediacaran är mindre klar. Det potentiella bidraget från detritalt, metamorfiskt förändrat organiskt material till prekambriska sedimentära successioner med låg TOC-halt är en möjlig mekanism för skillnader mellan olika platser i δ13CTOC64. TOC-innehållet i våra prover är dock, även om det är lågt, generellt sett större än 0,10 viktprocent, vilket avslöjar inga samband mellan TOC-innehåll och δ13C-värden. Dessutom innehåller våra prover termiskt omoget organiskt material och biomarkörlipidmönster som inte stämmer överens med en huvudsakligen alloktonisk kolkälla. Mer 13C-berikade δ13CTOC-signaturer kan vara resultatet av alternativa mekanismer för kolassimilation. Mekanismer för kolkoncentration eller aktivt bikarbonatupptag av prokaryoter kan resultera i mindre värden för εp och högre δ13CTOC-värden67,68. Uppkomsten av biota från Ediacaran kan ha utökat de marina näringsvägarna avsevärt och stimulerat nya vägar för mikrobiell heterotrofi, inklusive möjliga bidrag från komplexa kolcykler inom bentiska mikrobiella mattor. Med ytterligare hänsyn till den potentiellt viktiga rollen för löst organiskt material69 som kolkälla återspeglar bredden av δ13CTOC-värden sannolikt en rad biogeokemiska kolcykelprocesser som kan vara unika för den evolverande marina miljön i Ediacaran och som kan vara relaterade till ett fenomen för vilket vi inte har några representativa moderna analoger.
Antika sammansättningar av lipidbiomarkörer och indikatorer med stabila isotoper för bentisk, mikrobiell mattaproduktion tillsammans med fixerad kvävebegränsning har hittats i de tidiga triassiska bergarterna från södra Kina70. I efterdyningarna av massutdöendet i slutet av Perm vid Meishan fanns det en stor topp i hopan/steranförhållandena (upp till ca 60) tillsammans med en stark förskjutning av Norg-isotopsignaturen till värden mellan 0 och -2‰, vilket stämmer överens med bakteriell diazotrofi. Detta är förknippat med en hög signal av 2-metylhopaner (2-metylhopanindex upp till 33 %) och distinkta metylalkaner som pekar på spridning av bentiska mikrobiella mattor. Vi observerar inte en liknande trend i kväveisotopsignatur eller biomarkörmönster som uppenbarligen skulle peka på en betydande tillförsel av mikrobiella mattor inom vår datamängd från Ediacaran, även om ett singulärt 15N-förbrukat värde hittades i våra prover (tabell 1). Därför är den starka bakteriesignalen som observerades i våra Baltica-prover sannolikt inte främst en signatur av bentiska mikrobiella mattor.
Paleoenvironmental sustenance of Ediacara biota vs. Demosponges
De uppenbara oligotrofa förhållandena över de epikratoniska och kontinentala marginalbassängerna i Baltica, när den drev från höga till låga latituder från sent Ediacaran till tidigt Kambrium, kan vara förknippade med antingen begränsad advektion av relativt näringsrika djupa vatten eller utarmning av näringsämnen, till följd av assimilering och rensning under transport och avlagring över dessa breda, grunt marina epikontinentala bassänger (Fig. 3). Dessa bassänger var episodiskt isolerade från haven och utvecklade hypersalinitet (t.ex. under Redkino-tiden) och bräckta förhållanden (t.ex. under Kotlin-tiden71). Långsiktig tektonisk stabilitet resulterade i en topografi med låg relief i Baltica i slutet av Ediacaran, som var mycket känslig för översvämningar och ineffektiv tillförsel av fosfor som härrör från vittring. I motsats till Baltica avslöjade biomarkörstudier av strata från Huqf Supergroup i Oman ett eutroft ekosystem, rikt på mikroalger30,31,59, men utan Ediacara-biota, till och med i omfattande utbrott i Omanbergen och Huqf-regionen, i de inre och yttre skelfilterna3. Paleogeografin i södra Omans saltbassäng har rekonstruerats för den sena neoproterozoiska tiden vid ~13° från ekvatorn på södra halvklotet, vilket i stort sett liknar Baltica72-paleolatiden.
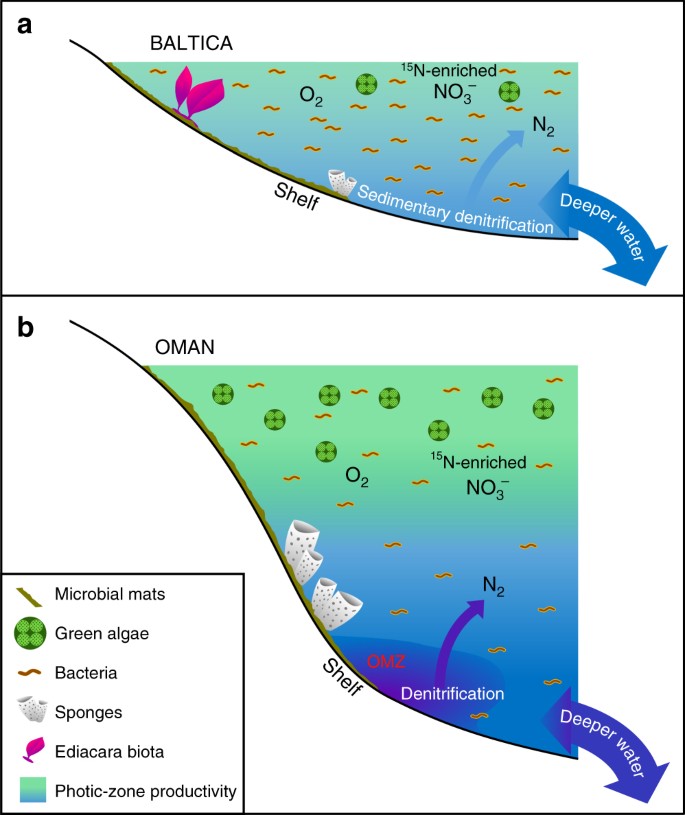
Större skillnader i havsmiljöer med låg produktivitet jämfört med produktiva Ediacaran-miljöer. Schematiska diagram visas för en omfattande oligotrof och ytlig marin epikontinental bassängmarginal i Baltica som ofta dominerades av bakteriell produktivitet där Ediacara mjukkroppsfauna blomstrade och där denitrifikation och anammox troligen var begränsade till sediment; och b eutrofa och djupare marina shelfmiljöer i södra Omans saltbassäng, där grönalger frodades som primärproducenter och demosvamparna var rikliga, men Ediacara-faunan med mjuka kroppar var inte framträdande (Ediacara-biota-fossil saknas också i korrelativa ediacariska utbrott i norra Oman). På produktiva kontinentala marginaler förekom troligen denitrifikation och anammox i både vattenpelaren och sedimenten
Det finns inga belägg för ihållande och omfattande anoxi på och runt de grunda kontinentala marginalerna i Baltica under den sena Ediacaran11, och vi föreslår att de oligotrofa förhållanden som beskrivs här orsakades av ineffektiva näringsflöden från land och djupvatten till dessa miljöer, som i stort sett liknar dem i oligotrofa miljöer i moderna havssystem. En geokemisk undersökning av spårelement i Utkina Zavod och intilliggande borrkärnor från S:t Petersburg-området tyder faktiskt på att våra prover avlagrades under oxiska förhållanden73. Denna tolkning stöds oberoende av våra data från de utökade hopanfördelningarna (C31-C35) som slutar kraftigt i abundans över C31-föreningar med ökande kolnummer, vilket är karakteristiskt för nedbrytning av sidokedjor av bakteriohopanpolyoler under diagenes under oxiska förhållanden29, och även av låga väteindex, som mäts genom Rock-Eval-pyrolys (alla lägre än 230 mg/g TOC, och många är lägre än 100 mg/g TOC) för dessa omogna prover på grund av bildandet av rekalcitrant kerogen via oxidativ nedbrytning och rekondensering av lipidfattig primärbiomassa i lokalt oxiska miljöer (tabell 1). De Ediacariska flercelliga organismer som existerade i dessa grunda marina miljöer måste ha haft tillräckligt med organiska substrat för heterotrofi för att tillgodose sina näringsbehov tillsammans med tillräckligt med syre och andra näringsämnen för att upprätthålla sin ämnesomsättning. De epikontinentala bassängerna i Baltica var sannolikt mer varaktigt oxiska än de högproduktiva miljöer som omgärdade syreminimumzoner, t.ex. på den mellersta till yttre shelfen i South Oman Salt Basin, där andning av riklig planktonbiomassa skulle ha upprätthållit lägre löst syre under den photiska zonen.
Ediacara biota och andra flercelliga organismer som lever utanför Baltica skulle ha varit tvungna att hantera förändrade näringskällor (bakteriella vs. eukaryotiska) när näringsflödet varierade med tiden, inklusive små celler och organiskt detritus i epikontinentala bassänger där bakterier var de dominerande primärproducenterna (fig. 3). Moderna korallrevssamhällen överlever i tropiska oligotrofa miljöer på grund av effektiv återvinning av näringsämnen, inklusive generering av ett flöde av löst organiskt material (DOM) som bidrar till att upprätthålla faunistisk heterotrofi i revets ekosystem74. Etableringen av en marin trofisk struktur med eukaryotiska flercelliga organismer som livnär sig på organiskt detritus måste ha skett efter den globala miljöexpansionen av eukaryoter i olika marina miljöer, som ägde rum under tonisk-kryogeniska intervallet (ca. 800-635 Ma), vilket kan bedömas utifrån biomarkörer15. Vi kan då förvänta oss att se bevis för betydande skillnader i den marina samhälls- och trofiska strukturen från plats till plats under den sena Ediacaranperioden, där den lokala näringsbalansen väljer eukaryotrika eller eukaryotfattiga mikrobiella samhällen och där den progressiva expansionen av flercelliga organismer tillför ytterligare en dimension av komplexitet på organism- och samhällsnivå. Betydande regionala kontraster i tillgången på fosfat och andra näringsämnen i shelfmiljöer är också en förväntad konsekvens av en heterogen global marin redoxstruktur för det sena Ediacaran, före syresättningen av djuphavet38.
Dominansen av picoplanktonbakteriell produktivitet och tillhörande nedbrytningsprodukter av löst organiskt material (DOM) i de epikontinentala bassängerna i Baltica skulle kunna ha upprätthållit ett mikrobiellt kretsloppsekosystem parallellt med den konventionella trofiska strukturen som bygger på omkring större planktonproducenter50. Detta kan ha gynnat olika former av heterotrofi, inklusive suspensionsfödning och eventuellt osmotrofi, som en livskraftig födostrategi för vissa rangeomorfer, svampar och andra flercelliga organismer från slutet av Ediacaran18 i samband med framväxande aktiv (rörlig) heterotrofi19,20. Näringsbegränsade akvatiska system domineras i regel ofta av små encelliga växtplankton och heterotrofa plankton, där bakterier konkurrerar ut eukaryoter och upprätthåller en låg nettobiomassa i oligotrofa marina miljöer i det moderna havet49,51. DOM är en viktig källa till organiska näringsämnen och styr ofta produktiviteten och nettobiomassan i moderna oligotrofa tropiska hav, men det gäller även kväve- och fosforklimatering75. På samma sätt var DOM sannolikt också ett viktigt substrat för att upprätthålla heterotrofa bakterier och ett mikrobiellt kretslopp i gamla oligotrofa miljöer. Bentiska mikrobiella mattor kan ha varit en del av denna bakteriellt dominerade näringsväv, och mattor har använts för att förbättra bevarandet av Ediacara-biota i marina miljöer7. Medan måttliga värden av 2-metylhopanindex hittades för en delmängd av våra prover (4-10 %, tabell 1), mycket låga förekomster av metylalkaner i förhållande till n-alkaner (fig. 1) och endast spår och sporadiska förekomster av karotenoider tyder på att mikrobiella mattor inte dominerade den primära produktiviteten76 , och att planktoniska bakterier och, på motsvarande sätt, deras nedbrytningsprodukter av DOM blomstrade. De generellt låga värdena för väteindex (tabell 1; särskilt för Redkino-proverna), trots den låga termiska mognaden hos strata och biomarkörsammansättningar, är också förenliga med lipidfattig organisk tillförsel i huvudsakligen oxiska, grunda marina marginella miljöer. Förekomsten av mikrobiella mattor, som endast bedöms utifrån sedimentologiska texturer, säger oss inte något om balansen mellan eukaryoter (mikroalger) och bakterier i vilket fall som helst, och de kan inte heller begränsa det relativa bidraget från mikrobiellt plankton till den totala primärproduktiviteten och till att upprätthålla näringsväven. Mikrobiella mattsamhällen som ofta innehåller rikligt med eukaryoter och Ara-gruppens karbonater från södra Oman med framträdande trombolitiska och krusiga laminitfacies är ett bra exempel på sena neoproterozoiska miljöer med betydande bidrag från mikrobiella mattor, som ger rikliga steransignaler på grund av ett stort bidrag från mikroalger30,59.
Den extremt låga nivån av 24-ipc-biomarkörer antyder att även om demosponger ibland förekom, så var de sparsamma i dessa miljöer. Detta kan tyda på färre möjligheter för mindre filterätande djur i konkurrens med Ediacara-biota i dessa miljöer med låg produktivitet, otillräcklig motståndskraft hos demosvamparna mot mer energirika grunda marina förhållanden ovanför vågbasen vid vackert väder, eller bättre anpassningsförmåga hos svamparna till syrefattiga förhållanden som dynamiskt upprätthålls under den photiska zonen i eutrofa miljöer (fig. 3). Totalt sett upptäcktes 24-ipc inte i de flesta proverna från Redkinohorisonten, som också generellt sett har de högsta H/St-förhållandena i vår provuppsättning, vilket kanske tyder på en ekologisk förändring vid Kotlinhorisontens avlagringstid. Alternativt är det också rimligt att svampar bebodde dessa miljöer, men inte producerade dessa diagnostiska steroidbiomarkörer i överflöd, även om detta verkar mindre troligt eftersom 24-ipc-steran är bland de vanligaste C30-steranföreningarna som påvisas i ediacaranska strata och oljor30,3133,59,60 och också kan påvisas i en delmängd av våra prover (tabell 1).
Och även om de ihållande oligotrofa marina miljöer som våra data tyder på representerar lokala förhållanden i de ediakariska oceanerna, var de sannolikt inte ovanliga för prekambriska grunda marina havsvägar (fig. 3). Den stora majoriteten av tidigare studier av biomarkörer från Ediacaran har utförts på organiskt rika sedimentära bergarter som avsatts i eutrofa miljöer och deras petroleumprodukter, som i allmänhet ger biomarkörsamlingar som är förenliga med ett betydande bidrag från mikroalger som källa44. Våra resultat understryker vikten av att studera en större variation av avlagringsmiljöer, inklusive organisk-materialfattiga lager med lämplig termisk mognad och olika litologier15,3034,59, för att få en mer exakt bild av omfattningen av heterogeniteten i marin kemi och ekologi från plats till plats. Trots progressiv havsventilation och ökad kemisk vittring och näringstillförsel under Rodinias upplösning och under hela Ediacaranperioden77,78, upprätthöll havets heterogenitet en mängd olika marinkemiska förhållanden, inklusive näringsfattiga, men beboeliga miljöer som främjade metazoers anpassning, konkurrens och evolution inom det globala havssystemet (fig. 3). Eutrofa marina hyllor var värd för demosvampar, men saknade ofta Ediacara-biota, med South Oman Salt Basin (och de korrelativa tjocka Ediacaranutskotten längre norrut i Oman) som ett framträdande exempel; tvärtom koloniserades grunda, oxiska och mindre produktiva epikontinentala havsvägar av Ediacara-biota i stället för av demosvampar, trots att DOM och annan organisk detritus fanns tillgängligt lokalt för födointag. Det är fortfarande osäkert om metaboliska krav eller selektivt tryck från miljön begränsade Ediacarans mjukkroppade flercelliga biota till dessa miljöer. Vår studie belyser dock att Ediacarans oligotrofa miljöer spelade en potentiellt avgörande roll för utvecklingen av makroskopiska flercelliga organismer och ekologin i de marina samhällena.