Maturitate termică excepțional de scăzută a stratelor
Maturitatea termică este cel mai important factor care influențează conservarea biomarkerilor sedimentari antici, deoarece moleculele organice sunt susceptibile de alterare structurală și stereochimică în timpul îngropării progresive și odată cu creșterea stresului termic29. Rocile precambriene cu o maturitate termică adecvată (maturitate de fereastră petrolieră sau mai mică) sunt condiții prealabile pentru conservarea unor ansambluri robuste și primare de lipide biomarker. Rapoartele multiple de stereoizomeri ai biomarkerilor hopan și steran, împreună cu dovezi independente din valorile Tmax scăzute (majoritatea în intervalul 417-433 °C, cu o medie de 426 °C) din piroliza Rock-Eval, indică faptul că rocile din acest studiu reprezintă de departe cele mai imature termic roci ediacariene analizate până în prezent cu ajutorul metodelor geochimice organice de ultimă generație (tabelul 1). Deși rocile noastre au suferit diageneză sedimentară, ele nu au trecut semnificativ în fereastra petrolieră, cu excepția probelor de afloriment 16PL ușor mai mature din bazinul Podillya din Moldova, care sunt încă adecvate pentru analiză (maturitate timpurie spre medie a ferestrei petrolifere și niciun semn evident de contaminanți organici). Cea mai mare parte a setului de atunci nu a fost probabil expusă la temperaturi de înmormântare care să depășească 50 °C, astfel că probele noastre sunt foarte imature. Alte linii de dovezi moleculare, care susțin o maturitate termică și o sinogenitate scăzută, includ (i) o dominanță a alcanilor policiclici biomarker față de n-alcani în extractele de rocă (Fig. 2), (ii) supraviețuirea unor cantități detectabile de 17β,21β(H)-hopani rezolvabili din hopani mai abundenți care posedă configurații stereochimice stabile 17β,21α(H) și 17α,21β(H) (Tabelul 1, Figurile suplimentare. 1 și 2), (iii) o preferință perceptibilă impară față de pară în rândul n-alcanilor din intervalul C22-C27 (Fig. 2), deoarece n-alcanii prezintă o preferință în funcție de numărul de carbon numai înainte de catageneză38 și (iv) generarea de biomarkeri de hopani și sterani imaturi din punct de vedere termic din faza kerogenică (insolubilă) cu ajutorul hidropirrolizei catalitice (Fig. 2). Maturitatea termică scăzută a materiei organice sedimentare din probele noastre este în concordanță cu indicatorii de alterare sedimentară publicați anterior, inclusiv cu indicii de alterare a conodontului și a acritarului, și cu studiile anterioare Rock-Eval privind piroliza și biomarkerii din rocile sedimentare ediacariene și paleozoice din Baltica36,39,40, precum și cu mineralogia argilei41. În mod critic, în mediile în care maturitatea termică este atât de scăzută, mecanismele de alterare semnificativă a semnalelor primare δ15N și δ13C sunt în mare parte absente42.

Distribuția hidrocarburilor alifatice extractibile pentru o probă reprezentativă. a Cromatograma ionică totală (TIC) pentru hidrocarburi alifatice extractibile pentru Lugovoe #13-73 m din orizontul Redkino. Seria n-alcanilor, pristanul (Pr), fitanul (Ph) și hopanii C27-C34 (indicați prin numărul total de atomi de carbon și stereochimia lor la C-17, C-21 și C-22, de exemplu, C31αβR) sunt etichetați; ** indică hopenii C29 și C30. Se remarcă dominanța abundenței hopenilor față de alte clase de compuși alcani. b Cromatograma parțială de ioni 85-Da arată dominanța seriei de n-alcani, prezentând o preferință perceptibilă a numerelor de carbon impare față de cele pare în intervalul C22-C27, față de metilalcani
Programe de asamblare a biomarkerilor lipidici
Un raport de abundență al hopanilor majori (C27-C35) față de steranii majori (C27-C29) este adesea folosit pentru a evalua echilibrul dintre aporturile de organisme sursă bacteriene și eucariote în ecosistemul acvatic antic. Hopanii sunt fosile moleculare derivate din hopanoide, care sunt lipide de membrană celulară sintetizate de o mare varietate de grupuri bacteriene. În mod similar, steranii sunt derivați din precursori de steroli, care sunt produși aproape exclusiv de eucariote43. Rapoartele hopano/esterani (H/St) pentru probele noastre acoperă o gamă surprinzător de largă de valori, de la 1,6 la 119,2 (tabelul 1). Pentru context, raporturile H/St din rocile și uleiurile neoproterozoice bogate în substanțe organice se încadrează de obicei într-un interval îngust de la 0,5 la 2,030,31,32,33. Deși există o diferență clară între valorile din orizontul Kotlin mai tânăr (H/St mediu de 8,9) și orizontul Redkino mai vechi (H/St mediu de 42,9), valorile pentru majoritatea probelor sunt neobișnuit de ridicate și sugerează contribuții anormal de ridicate ale bacteriilor.
Până la sfârșitul Neoproterozoicului, algele eucariote erau o componentă semnificativă din punct de vedere ecologic și producători majori în multe ecosisteme marine15,34,44. Discrepanța dintre contribuția importantă la nivel global a algelor eucariote la sfârșitul Neoproterozoicului și totuși nivelurile scăzute ale biomarkerilor de steran în aceste probe indică faptul că trebuie să existe un determinant local asupra abundenței eucariote. Cea mai parcimonioasă explicație pentru ratele extrem de ridicate de hopan/steran, alături de conținuturile scăzute de carbon organic total (TOC) și de indicii de hidrogen (HI) scăzuți găsiți pentru probele noastre (tabelul 1) este că aceste straturi au fost depuse în medii oligotrofe (adică puternic limitate de nutrienți), în care bacteriile au surclasat algele. În timp ce analogii moderni pentru mările epicratonice antice dezvoltate în timpul unui nivel ridicat al mării sunt greu de găsit, observații paralele ale unor rapoarte hopan/steran mai mari (cu până la un ordin de mărime) au fost găsite anterior pentru rocile sedimentare sărace în substanțe organice față de cele bogate în substanțe organice depuse în căile maritime epicontinentale din Ordovician-Silurian45.
În mod obișnuit, un conținut scăzut de COT poate fi cauzat de depunerea limitată a materiei organice în medii cu productivitate scăzută, de potențialul scăzut de conservare a materiei organice în coloana de apă sau în sedimente, sau de diluția cu un flux siliciclastic ridicat. În oceanele moderne, productivitatea este cel mai adesea limitată de nivelurile scăzute de nutrienți esențiali: azot, fosfor, siliciu și fier46. Azotul, fosforul și fierul sunt limitate în regiunile din largul oceanului, în care afluența, aportul de praf și scurgerile de coastă nu furnizează cantități suficiente de nutrienți. În mediile oligotrofe moderne, raportul dintre biomasa bacteriană și cea eucariotă este mai mare decât în mediile eutrofe sau mezotrofe. În mod interesant, probele de afloriment Podillya 16PL din orizontul Redkino din Moldova sunt luturi care conțin fosforită și dau rapoarte hopan/steran semnificativ mai mici (8,1:11,5) decât celelalte probe Redkino din tabelul 1 (deși sunt încă semnificativ mai mari decât cele găsite în Bazinul Salinic din sudul Omanului sau în sedimentele fanerozoice bogate în substanțe organice, care se încadrează de obicei într-un interval îngust, de 0,5 %.5-2,0), sugerând probabil că disponibilitatea crescută a fosfaților ar fi putut influența în mod favorabil raportul eucariote/bacterii găsit la nivel local (având în vedere că semnăturile izotopice ale azotului sunt în mare parte invariabile, a se vedea secțiunea următoare). Conținutul de fosfor (P), precum și raporturile P/Fetotal și P/Al, sunt de altfel, în general, scăzute pentru sedimentele ediacariene târzii din Podillya, Ucraina și Estonia47; cu excepția acestui nivel stratigrafic marcat cu noduli de fosforită. În mod similar, niveluri scăzute (~0,01 până la ~0,1 % în greutate) de P în rocile siliciclastice Kotlin și Redkino au fost raportate de la o carotă de foraj de la marginea nord-estică a Platformei Est-Europene11.
Topografia largă și puțin adâncă din mările epicontinentale din Baltica ar fi putut susține limitări ale fosforului sau ale altor nutrienți în mediile marginale, datorită precipitării authigenice a fosfatului cu minerale de fier în apele de suprafață oxicale și a sechestrării unei serii de elemente de către sedimentele de platou48. În cazul în care fosfatul a fost un nutrient limitativ, creșterea fosforului biodisponibil ar fi sporit producția primară locală și ar fi oferit condiții de creștere mai favorabile pentru fitoplanctonul unicelular de dimensiuni mai mari49,50,51. În ceea ce privește sistemul oceanic modern, picocyanobacteriile marine, Prochlorocococcus și Synechococcus, sunt recunoscute ca fiind cele care domină numărul de celule și biomasa fitoplanctonului în mediile oligotrofe ale oceanelor tropicale și subtropicale, inclusiv în regiunile oligotrofe limitate de fosfați din apele de suprafață ale oceanelor52. Prochlorococcus și SAR11 heterotrofic (Pelagibacter) prosperă datorită unei varietăți de adaptări, inclusiv costuri energetice reduse în virtutea unor genomuri mici și a unor rate de replicare scăzute, un raport suprafață-volum mai mare datorită dimensiunilor mai mici ale celulelor și funcții suplimentare de absorbție celulară pentru a maximiza utilizarea nutrienților53. Multe bacterii sunt, de asemenea, capabile să înlocuiască nutrienții cu abundență redusă, de exemplu, prin utilizarea sulfolipidelor în locul lipidelor purtătoare de fosfor în mediile cu deficit de P54 sau prin utilizarea de substraturi alternative, de exemplu, aprovizionarea cu azot din N2 atmosferic prin fixarea azotului55 , pentru a atenua stresul nutritiv. Un studiu recent privind supraviețuirea bacterioplanctonului marin în medii oligotrofe56 , unde fosfatul disponibil este limitat, sugerează un rol important pentru metabolismul polifosfaților în oligotrofii marini. Eucariotele selecționate care concurează în medii oligotrofe sunt, de obicei, picoeucariote mici, care își pot suplimenta cerințele nutriționale prin mixotrofie57. Raportul dintre biomasa de picoeucariote și cianobacterii tinde să crească în cazul unei aprovizionări sporite cu nutrienți49. Posibila influență a fosforului, precum și a altor nutrienți biolimitanți, în moderarea productivității primare și a structurii comunităților marine în căile maritime epeirice ediacariene necesită investigații suplimentare.
Fără excepție, abundența steranilor C29 este mai mare decât cea a steranilor C27 sau C28 corespunzători pentru toate locațiile noastre (tabelul 1). O predominanță a steranilor C29 față de steranii C27 și C28 indică probabil o predominanță a algelor verzi în cadrul comunității fitoplanctonice eucariote44,58. Această caracteristică a fost observată în majoritatea studiilor anterioare privind biomarkerii din Ediacaran15,30,31,31,32,33,33,34,35,36,44. În mod special, distribuția steranului C30 în mai multe probe din fiecare carotă de foraj din Orizontul Kotlin conține cantități scăzute, dar detectabile, ale biomarkerului steran de demosponge cunoscut sub numele de 24-isopropilcolestan (24-ipc)59,60. În total, steranii 24-ipc au fost raportați în eșantioane de rocă și de petrol care datează încă din Criogenian (>635 Ma în urmă) în Bazinul Sărat din sudul Omanului și reprezintă cea mai veche dovadă de biomarker lipidic pentru metazoare59,60. Biomarkerii 24-ipc din probele noastre au fost fie cu aproximativ un ordin de mărime mai mici în abundență (în raport cu raportul total al steranilor C27 la C30, aceștia au fost de numai 0,06-0,61%; media = 0,22%) în comparație cu rocile și uleiurile din Ediacaran din Omanul de Sud (1,7% în medie59), fie au fost sub limitele de detecție din cauza abundenței neglijabile pentru majoritatea probelor.
Raporturile izotopice ale azotului și carbonului organic
Izotopii de azot pot ajuta la discernerea echilibrului relativ în ciclul azotului și a gradului în care fie fixarea azotului, fie denitrificarea incompletă au fost căi semnificative pentru a influența echilibrul nutrienților disponibili pentru comunitățile marine. Atunci când bacteriile diazotrofe fixează azotul molecular din cauza lipsei de azot fixat în coloana de apă, acest lucru poate produce valori izotopice ale azotului masiv sedimentar aproape de 0‰61. Acest lucru este în contrast cu semnăturile izotopice pozitive ale azotului (în intervalul +2 până la +10‰), cu un mod de +4 până la +6‰, găsite pentru sedimentele marine neoproterozoice depuse în ceea ce se consideră a fi condiții bogate în nitrați, în care nitratul a suferit doar o denitrificare parțială62. Valorile izotopilor de azot pentru toate probele noastre, cu excepția unei valori aberante (tabelul 1), acoperă un interval pozitiv limitat de la +3,5 la +6,5‰, care se suprapune cu modul pentru rocile sedimentare bogate în materie organică din Neoproterozoicul târziu62. Constrânse de indicii de redox care sugerează că au predominat condițiile oxicale din coloana de apă, datele noastre implică faptul că nitrații au dominat fondul de azot anorganic dizolvat. Gama de valori δ15N sugerează, de asemenea, că fixarea N2 nu a fost principalul mod de achiziție a azotului pentru producătorii primari. Mai degrabă, ciclul azotului a fost probabil dominat de reciclarea coloanei de apă, iar N organic a fost supus oxidării cantitative la nitrați, așa cum se întâmplă în mediile marine moderne, proximale. Denitrificarea bentonică și îngroparea azotului organic ar fi fost rezervoarele primare pentru azotul anorganic dizolvat, iar gama de valori δ15N sugerează doar un rol limitat pentru denitrificarea incompletă din coloana de apă, care are ca rezultat, de obicei, o îmbogățire semnificativă în 15N63. Prin urmare, variabilitatea limitată a valorilor δ15N reflectă probabil δ15N al nitraților aduși pe platformă, cu o influență minoră a reducerii limitate a nitraților din coloana de apă și a fixării N2. Fără dovezi puternice δ15N pentru fixarea azotului, presupunem că azotul nu a fost principalul nutrient biolimitant în bazinele epicontinentale din Baltica. Este posibil ca izolarea de sursele fluviale și eoliene de fosfor și condițiile oxicale din bazinele epicontinentale largi, de mică adâncime marină, să fi favorizat eliminarea fosforitei authigenice și a fazelor purtătoare de urme de metal, rezultând o limitare a nutrienților care a constrâns creșterea și producția de celule eucariote.
Valorile izotopice ale carbonului organic total (CTOC) variază de la -23,0 la -33,9‰, cele mai mari diferențe fiind observate între diferitele locații ale carotelor de foraj. Îmbogățirea relativă a 13C la raporturi izotopice mai mari de cca. -28‰ contrastează cu datele din stratele contemporane depuse în mediile eutrofe, deschise-marine din Oman30,31. Cu toate acestea, intervalul pe care îl raportăm este, în general, în concordanță cu intervalul δ13CTOC pentru alte locații din Baltica depozitate în aceeași perioadă de timp11. Diferența dintre Baltica și Oman ar putea evidenția faptul că valorile δ13CTOC nu reflectă exclusiv o schimbare seculară uniformă a ciclului carbonului în acest interval de timp31,64. Mai degrabă, intervalul δ13CTOC de la Baltica poate reflecta, în parte, ecologia microbiană dominată de bacterii, sugerată de raporturile unice ale biomarkerilor lipidici. Dimensiunea mică a celulelor, raportul mare dintre suprafață și volum și ratele de creștere lentă în condiții oligotrofe pot crește magnitudinea fracționării în timpul autotrofiei (εp)65,66, rezultând în valori δ13C scăzute.
Mecanismul pentru semnăturile izotopice mai îmbogățite în 13C din intervalul raportat11 pentru succesiunea de la Baltica de la sfârșitul Ediacarului este mai puțin clar. Contribuția potențială a materiei organice detritale, alterate metamorfic, la succesiunile sedimentare precambriene cu conținut scăzut de COT este un mecanism posibil pentru disparitățile dintre situri în δ13CTOC64. Cu toate acestea, conținutul de COT al probelor noastre, deși scăzut, este în general mai mare de 0,10 % în greutate, ceea ce nu relevă nicio relație între conținutul de COT și valorile δ13C. Mai mult, eșantioanele noastre conțin materie organică imatură din punct de vedere termic și modele de lipide biomarker care nu sunt în concordanță cu o sursă de carbon în principal alohtonă. Semnăturile δ13CTOC mai îmbogățite cu 13C pot fi rezultatul unor mecanisme alternative de asimilare a carbonului. Mecanismele de concentrare a carbonului sau absorbția activă a bicarbonatului de către procariote pot avea ca rezultat valori mai mici pentru εp și valori δ13CTOC mai mari67,68. Este posibil ca apariția biotei ediacariene să fi extins în mod semnificativ rețelele trofice marine și să fi stimulat noi căi de heterotrofie microbiană, inclusiv posibilele contribuții ale ciclului complex al carbonului în cadrul covoarelor microbiene bentice. Luând în considerare, în plus, rolul potențial important al materiei organice dizolvate69 ca sursă de carbon, amploarea valorilor δ13CTOC reflectă probabil o serie de procese biogeochimice de reciclare a carbonului care ar putea fi unice pentru mediul marin ediacaran în evoluție și ar putea fi legate de un fenomen pentru care nu avem analogi moderni reprezentativi.
În rocile triasice timpurii din sudul Chinei au fost descoperite ansambluri de biomarkeri lipidici antici și indicatori de izotopi stabili pentru producția bentonică, microbiană de covor în tandem cu limitarea azotului fixat70. În urma extincției în masă de la sfârșitul Permianului de la Meishan, a existat o creștere mare a rapoartelor hopan/steran (până la aproximativ 60), însoțită de o schimbare puternică a semnăturii izotopice Norg la valori cuprinse între 0 și -2‰, în concordanță cu diazotrofia bacteriană. Acest lucru este asociat cu un semnal ridicat de 2-metilhopani (indice de 2-metilhopani de până la 33%) și metilalcani distinctivi care indică proliferarea covoarelor microbiene bentice. Nu observăm o tendință similară în ceea ce privește semnătura izotopică a azotului sau modelele de biomarker care ar indica în mod evident o contribuție semnificativă a covorului microbian în cadrul setului nostru de date Ediacaran, deși în probele noastre a fost găsită o valoare singulară sărăcită de 15N (tabelul 1). Prin urmare, este probabil ca semnalul bacterian puternic observat în probele noastre de la Baltica să nu fie predominant o semnătură a covoarelor microbiene bentonice.
Susținerea paleoambientală a biotei Ediacara vs. Ediacara. demosponge
Condițiile aparent oligotrofe din bazinele epicratonice și de margine continentală ale Balticii, pe măsură ce a derivat de la latitudini înalte la latitudini joase de la sfârșitul Ediacarului până la începutul Cambrianului, ar putea fi asociate fie cu advecția limitată a apelor de adâncime relativ bogate în nutrienți, fie cu epuizarea nutrienților, rezultată din asimilarea și curățarea în timpul transportului și depunerii în aceste bazine epicontinentale epicontinentale largi, de mică adâncime marină (Fig. 3). Aceste bazine au fost izolate episodic de oceane și au dezvoltat hipersalinitate (de exemplu, în timpul perioadei Redkino) și condiții salmastre (de exemplu, în timpul perioadei Kotlin71). Stabilitatea tectonică pe termen lung a dus la o topografie cu relief redus a Balticii Ediacariene târzii, foarte susceptibilă la inundații și la un aport ineficient de fosfor derivat din meteorizare. Spre deosebire de Baltica, studiile privind biomarkerii stratelor din Supergrupul Huqf din Oman au evidențiat un ecosistem eutrofic, bogat în microalge30,31,59, dar lipsit de biota Ediacara, chiar și în aflorimentele extinse din Munții Oman și din regiunea Huqf, în mediile interioare și exterioare ale platoului3. Paleogeografia bazinului salinic din sudul Omanului a fost reconstituită pentru Neoproterozoicul târziu la ~13° față de ecuator în emisfera sudică, în linii mari similară paleolatitudinii Baltica72.
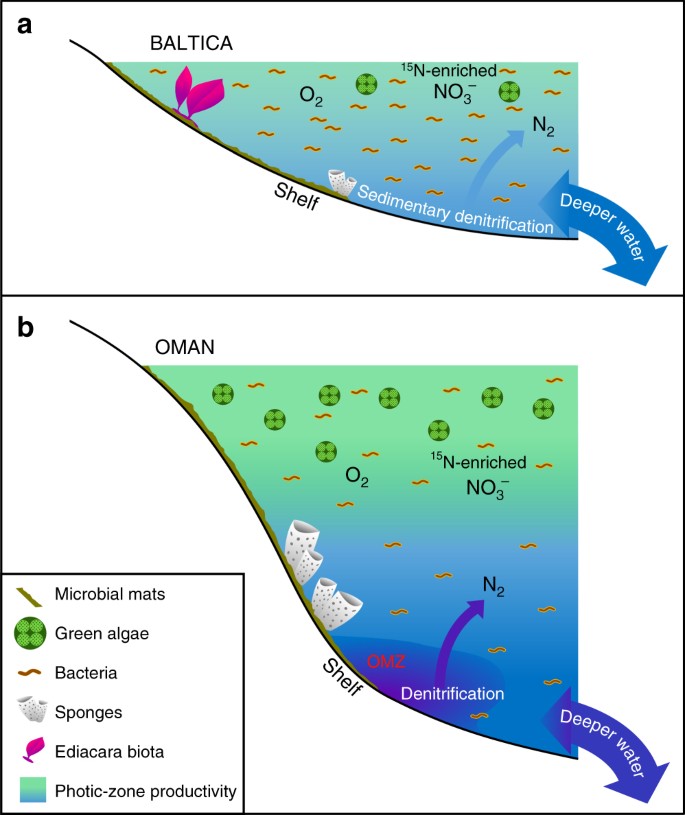
Diferențe majore între mediile marine Ediacariene cu productivitate scăzută și cele productive. Sunt prezentate diagrame schematice pentru un bazin epicontinental oligotrofic extins și de mică adâncime de la marginile bazinelor din Baltica, adesea dominat de productivitatea bacteriană, unde fauna cu corpuri moi Ediacara a înflorit și unde denitrificarea și anammoxul au fost probabil limitate la sedimente; și b. mediile de platou eutrofic și de mare adâncime din bazinul de sare din sudul Omanului, unde algele verzi au prosperat ca producători primari și demospongele au fost abundente, dar fauna de corpuri moi Ediacara nu a fost proeminentă (fosilele de biota Ediacara fiind, de asemenea, absente în aflorimentele Ediacariene corelate din nordul Omanului). Pe marginile continentale productive, denitrificarea și anammoxul au avut loc probabil atât în coloana de apă, cât și în sedimente
Nu există dovezi pentru o anoxie persistentă și extinsă pe și în jurul marginilor continentale de mică adâncime din Baltica în timpul Ediacaranului târziu11, și sugerăm că condițiile oligotrofe descrise aici au fost cauzate de fluxurile ineficiente de nutrienți tereștri și de adâncime către aceste medii, în linii mari similare cu cele din mediile oligotrofe din sistemele oceanice moderne. Într-adevăr, o investigație geochimică a oligoelementelor din Utkina Zavod și a carotelor de foraj adiacente din zona Sankt Petersburg sugerează că probele noastre au fost depuse în condiții oxicale73. Această interpretare este susținută în mod independent de datele noastre din distribuțiile extinse ale hopanilor (C31-C35), care se reduc brusc în abundență deasupra compușilor C31 odată cu creșterea numărului de carbon, ceea ce este caracteristic degradării lanțului lateral al bacteriohopanepoliolilor în timpul diagenezei în condiții oxicale29, și, de asemenea, de indicii cu conținut scăzut de hidrogen, măsurați prin piroliza Rock-Eval (toți sunt mai mici de 230 mg/g COT, iar mulți sunt sub 100 mg/g COT) pentru aceste probe imature, datorită formării de kerogen recalcitrant prin degradarea oxidativă și recondensarea biomasei primare sărace în lipide în medii local oxicale (tabelul 1). Organismele pluricelulare ediacariene care au existat în aceste medii marine de mică adâncime trebuie să fi avut suficiente substraturi organice pentru heterotrofie pentru a-și satisface nevoile de hrană, împreună cu suficient oxigen și alți nutrienți pentru a-și susține metabolismul. Bazinele epicontinentale din Baltica au fost probabil mai persistent oxic decât mediile foarte productive care mărginesc zonele cu oxigen minim, cum ar fi pe platoul mijlociu spre exterior al bazinului de sare din sudul Omanului, unde respirația biomasei planctonice abundente ar fi menținut un nivel mai scăzut de oxigen dizolvat sub zona fotică.
Biota Ediacara și alte organisme pluricelulare care trăiesc în largul Balticii ar fi trebuit să facă față unor surse de hrană în schimbare (bacteriene vs. eucariote) pe măsură ce fluxurile de nutrienți variau în timp, inclusiv celule mici și detritus organic în bazinele epicontinentale unde bacteriile erau producătorii primari dominanți (Fig. 3). Comunitățile moderne de recif de corali supraviețuiesc în mediile oligotrofe tropicale datorită reciclării eficiente a nutrienților, inclusiv generării unui flux de materie organică dizolvată (DOM) care ajută la susținerea heterotrofiei faunei în cadrul ecosistemului de recif74. Stabilirea unei structuri trofice marine cu organisme eucariote multicelulare susținute prin hrănirea cu detritus organic a trebuit să fie ulterioară expansiunii ecologice la scară globală a eucariotelor în diverse medii marine, care a avut loc în intervalul Tonian-Criogenian (cca. 800-635 Ma), așa cum reiese din înregistrările de biomarker15. Ne-am putea aștepta atunci să găsim dovezi ale unor diferențe semnificative în ceea ce privește comunitatea marină și structura trofică de la o localitate la alta în perioada Ediacaranului târziu, echilibrul nutritiv local selectând comunități microbiene bogate sau lipsite de eucariote, iar expansiunea progresivă a organismelor pluricelulare adăugând o altă dimensiune a complexității la nivel de organism și de comunitate. Contrastul regional semnificativ în ceea ce privește disponibilitatea fosfaților și a altor nutrienți în mediile de platou este, de asemenea, o consecință așteptată a unei structuri redox marine globale eterogene pentru Ediacaranul târziu, înainte de oxigenarea oceanului de adâncime38.
Dominanța productivității bacteriene a picoplanctonului și a produselor asociate de degradare a materiei organice dizolvate (DOM) în bazinele epicontinentale din Baltica ar fi putut susține un ecosistem microbian în buclă, în paralel cu structura trofică convențională bazată pe producători planctonici de dimensiuni mai mari50. Este posibil ca acest lucru să fi favorizat diferite moduri de heterotrofie, inclusiv hrănirea în suspensie și, posibil, osmotrofia, ca strategie viabilă de hrănire pentru unele rangeomorfe, bureți și alte organisme multicelulare de la sfârșitul Ediacarului18, în combinație cu heterotrofia activă (mobilă) emergentă19,20. Sistemele acvatice limitate de nutrienți, ca regulă generală, sunt adesea dominate de fitoplancton unicelular de mici dimensiuni și de plancton heterotrof, cu bacterii care depășesc eucariotele și care susțin o biomasă netă scăzută în mediile marine oligotrofe ale oceanului modern49,51. DOM este o sursă importantă de nutrienți organici și controlează adesea productivitatea și biomasa netă în mările tropicale oligotrofe moderne, dar la fel este și co-limitarea azotului și a fosforului75. În mod similar, DOM a fost, de asemenea, probabil, un substrat important pentru susținerea bacteriilor heterotrofe și o buclă microbiană în mediile oligotrofe antice. Este posibil ca rogojini microbiene bentonice să fi fost o componentă a acestei rețele alimentare dominate de bacterii, iar terenurile de rogojini au fost implicate pentru a spori conservarea biotei Ediacara în medii marine7. Deși s-au găsit valori moderate ale indicelui de 2-metilhopane pentru un subset de probe (4-10%, tabelul 1), abundența foarte scăzută a metilalcanilor în raport cu n-alcanii (Fig. 1) și doar urme și prezențe sporadice de carotenoizi sugerează că covorașele microbiene nu au dominat productivitatea primară76 și că bacteriile planctonice și, în mod corespunzător, produsele lor de descompunere a DOM au prosperat. Valorile în general scăzute ale indicelui de hidrogen (tabelul 1; în special pentru probele Redkino), în ciuda maturității termice scăzute a straturilor și a ansamblurilor de biomarkeri, sunt, de asemenea, în concordanță cu aportul organic sărac în lipide în medii marginale, în principal oxic, de mică adâncime. Prezența maturilor microbiene, măsurată doar din texturile sedimentologice, nu ne spune în niciun caz despre echilibrul dintre eucariote (microalge) și bacterii, și nici nu poate constrânge contribuția relativă a planctonului microbian la productivitatea primară globală și la susținerea rețelelor trofice. Comunitățile de covorașe microbiene care conțin adesea eucariote abundente, iar carbonații din Grupul Ara din Omanul de Sud, cu faciesuri trombolitice proeminente și laminite încrețite, sunt un bun exemplu de medii neoproterozoice târzii cu o contribuție semnificativă a covorașelor microbiene, care dau semnale steriene abundente datorită unei contribuții mari a microalgelor30,59.
Nivelul extrem de scăzut al biomarkerilor 24-ipc sugerează că, deși demospongele au fost uneori prezente, acestea au fost rare în aceste medii. Acest lucru ar putea indica mai puține oportunități pentru animalele mai mici care se hrănesc prin filtrare în competiție cu biota Ediacara în aceste medii cu productivitate scăzută, o rezistență insuficientă a demospongei față de condițiile marine de mică adâncime mai energetice de deasupra bazei de valuri de vreme bună sau o mai bună adaptabilitate a bureților la condițiile cu conținut scăzut de oxigen menținute în mod dinamic sub zona fotică în mediile eutrofice (Fig. 3). În total, 24-ipc nu a fost detectat în majoritatea eșantioanelor din Orizontul Redkino, care au, de asemenea, în general, cele mai mari rapoarte H/St din setul nostru de eșantioane, ceea ce indică, probabil, o schimbare ecologică până la momentul depunerii din Orizontul Kotlin. Alternativ, este, de asemenea, plauzibil ca bureții să fi locuit în aceste medii, dar să nu fi produs acești biomarkeri steroizi de diagnosticare în abundență, deși acest lucru pare mai puțin probabil, deoarece steranul 24-ipc se numără printre cei mai frecvent detectați compuși steranici C30 în stratele și uleiurile ediacariene30,3133,59,60 și este, de asemenea, detectabil într-un subset al probelor noastre (tabelul 1).
În timp ce mediile marine oligotrofe persistente sugerate de datele noastre reprezintă condiții localizate în oceanele ediacariene, acestea nu au fost probabil neobișnuite pentru căile maritime precambriene de mică adâncime (Fig. 3). Marea majoritate a studiilor anterioare privind biomarkerii din Ediacaran au fost efectuate pe roci sedimentare bogate în substanțe organice depuse în medii eutrofe și pe produsele lor petroliere, care, în general, produc ansambluri de biomarkerii în concordanță cu o contribuție semnificativă a sursei microalgale44. Rezultatele noastre evidențiază importanța studierii unei varietăți mai mari de medii de depunere, inclusiv a straturilor sărace în materie organică cu o maturitate termică adecvată și diferite litologii15,3034,59, pentru a obține o imagine mai precisă pentru scara eterogenității în chimia și ecologia marină de la un loc la altul. În pofida ventilației progresive a oceanului și a creșterii meteorizării chimice și a aprovizionării cu nutrienți în timpul dezmembrării Rodiniei și pe parcursul perioadei ediacariene77,78, eterogenitatea oceanului a menținut o varietate de condiții chimice marine, inclusiv medii sărace în nutrienți, dar locuibile, care au favorizat adaptarea, competiția și evoluția metazoarelor în cadrul sistemului oceanic global (Fig. 3). În timp ce platformele marine eutrofe au găzduit demosponge, dar adesea au lipsit biota Ediacara, bazinul de sare din sudul Omanului (și aflorimentele groase Ediacaran corelative mai la nord în Oman) fiind un exemplu proeminent; în mod contraintuitiv, căile maritime epicontinentale puțin adânci, oxicale și mai puțin productive au fost colonizate de biota Ediacara cu preferință față de demosponge, în ciuda faptului că DOM și alte detritusuri organice erau disponibile la nivel local pentru hrănire. Dacă cerințele metabolice sau presiunea selectivă a mediului au restricționat biota multicelulară cu corpuri moi din Ediacaran la aceste medii rămâne incert; cu toate acestea, studiul nostru evidențiază faptul că mediile oligotrofe din Ediacaran au jucat un rol potențial crucial în evoluția organismelor multicelulare macroscopice și în ecologia comunităților marine.
.